
Résumé
Le système nerveux central des mammifères adultes a longtemps été considéré comme une structure rigide, très précisément câblée, dont la réparation post-lésionnelle était impossible. Nous avons montré qu’il existe en fait un degré assez considérable de plasticité potentielle, dont les auteurs peuvent être les neurones eux-mêmes, mais surtout la névroglie, et les cellules souches précurseurs multipotentes. La moelle épinière constitue un modèle de choix sur lequel nous avons pu montrer au moyen de modèles lésionnels traumatiques et vasculaires qu’une intervention pharmacologique rapide peut réduire significativement l’étendue des lésions et leurs conséquences fonctionnelles. Par ailleurs, la repousse d’axones lésés peut être obtenue en modifiant la réactivité des astrocytes, principal élément de la réaction gliale. Enfin, des thérapies cellulaires substitutives (greffes neuronales) peuvent permettre de rétablir certaines fonctions réflexes dans une moelle totalement déconnectée des afférences cérébrales. L’ensemble de ces données ouvre des perspectives thérapeutiques nouvelles dans de nombreuses affections du système nerveux. Le syndrome de Turner est une dysgénésie gonado-somatique de phénotype féminin due à une monosomie plus ou moins complète d’un des chromosomes X, ce qui conduit à une haplo-insuffisance des gènes du développement situés dans la région pseudo-autosomale des gonosomes. La pratique plus systématique du caryotype a montré la prépondérance des mosaïques et la grande variabilité du phénotype turnérien en proportion du nombre de cellules 45,X. Les rares cas de monozygotisme discordant avec des mosaïques à distribution tissulaire variable permettent de comprendre que l’expression phénotypique est un effet de dosage génique. Ceci incite à étudier un deuxième tissu, comme les fibroblastes cutanés lorsqu’on constate chez une patiente avec ST une discordance entre le phénotype et le caryotype. Le mosaïcisme est donc la coexistence dans un organisme de cellules de constitution chromosomique ou génique différente, la proportion de cellules normales variable selon les tissus modulant le phénotype.
Summary
The central nervous system of adult mammals has been classically considered as structurally rigid, tightly wired, and unable to be repaired. We have shown that there exists a rather considerable degree of intrinsic plasticity due to the neurons themselves, but merely to glial cells and to multipotent stem cells. The spinal cord constitutes a good model on which we could demonstrate, with vascular and traumatic animal paradigms, that an early pharmacologic intervention could reduce significantly the extent of lesions and the subsequent functional deficit. Moreover, we showed that regeneration of severed central axons could occur, provided that the astrocytes’ component of the glial scar was modified. Finally, transplants of embryonic neurons were shown to repair the axonal circuitry below a sectioned cord, and to restore reflex functions. All these data point to unprecedented perspectives of efficient therapies in acute and chronic neurological diseases. Turner’s syndrome is a gonadosomatic dysgenesis of female phenotype due to a more or less complete monosomy of one of the X chromosomes leading to a haploinsufficiency of the development genes situated at the level of the pseudoautosomal region of the gonosomes. Further experience of the karyotype showed a preponderance of mosaics and considerable variability of Turner’s phenotype in proportion to the number of 45,X cells. The rare cases of monozygotism discordant with variable tissular distribution mosaics show that the phenotypic expression is a genic dosage effect. In patients with TS it would thus be of interest to study a second tissue such skin fibroblasts when a discordance is observed between the phenotype and the karyotype.
INTRODUCTION
Au cours des deux dernières décennies, le dogme établi par Cajal et ses élèves [1-4] de non régénération du système nerveux central des mammifères a été vivement contesté, entre autres par les travaux de A. Aguayo [5]. Il est apparu clairement que l’incapacité apparente des neurones centraux adultes de régénérer un axone sectionné n’était pas intrinsèque à ces cellules, mais dépendante d’un environnement cellulaire non permissif. Par ailleurs, les progrès de la neuropharmacologie, d’une part avec une meilleure identification des cibles [6] et de la biologie cellulaire et moléculaire, d’autre part avec l’élaboration de nouveaux outils expérimentaux et thérapeutiques [7-8], nous ont ouvert des axes de recherche sans précédents dans l’histoire des neurosciences. Sur ces bases, nous avons entrepris de mettre en œuvre une recherche multidisciplinaire centrée sur un modèle, la moelle épinière.
CHOIX DU MODÈLE
Notre choix a été guidé par la nécessité de disposer de paramètres anatomiques, biochimiques et fonctionnels cohérents, suffisamment sensibles pour pouvoir évaluer une éventuelle récupération, et que ce modèle corresponde à une pathologie humaine elle-même accessible à l’analyse. Les lésions traumatiques de la moelle épinière répondent à ces critères : l’anatomie de la moelle est bien connue, et aisément accessible à une micro-chirurgie réglée. Les entrées et sorties physiologiques, en particulier sensibilité et motricité, sont accessibles à une quantification rigoureuse. Enfin, l’anatomie même de la moelle se prête particulièrement bien à des prélèvements précis destinés à la biochimie.
ANATOMIE ET RÔLE CRITIQUE DES AFFÉRENCES MONOAMINERGIQUES
Les systèmes monoaminergiques centraux ne jouent pas de rôle direct dans les grandes fonctions nerveuses, mais ils interviennent de façon ubiquitaire dans le contrôle de ces fonctions [9]. La moelle épinière n’échappe pas à cette règle, avec une
concentration de noradrénaline, de sérotonine et, à un moindre degré, de dopamine, au niveau de 3 cibles essentielles à la physiologie médullaire : dans la corne postérieure, où s’articulent les afférences primaires avec les systèmes sensitifs ascendants, dans la colonne intermédio-latérale, où sont situés les neurones pré- ganglionnaires du système nerveux autonome, au niveau des motoneurones, dernière étape de la commande motrice.
La première étape de notre étude a consisté à analyser le développement pré et postnatal des afférences sérotoninergiques et noradrénergiques. Nous avons montré que le plus précoce est le système sérotonergique issu des noyaux du raphé caudal, dont les premiers axones envahissent la moelle au 15ème jour de gestation chez le rat [10]. La corne ventrale, puis la colonne intermédio-latérale, puis enfin la corne dorsale (au moment de la naissance) sont successivement envahies. Les premières synapses sont détectées au 17ème jour gestationnel dans la corne ventrale [10].
L’innervation noradrénergique est retardée d’un à deux jours suivant la région, et suit le même gradient ventro-dorsal [11-12].
Il est intéressant de remarquer que cette innervation précède de plusieurs jours la mise en place des systèmes moteurs et sensitifs. Il est probable — mais non prouvé directement, au moins au niveau de la moelle épinière — que les monoamines jouent un rôle trophique dans la période périnatale.
L’ultrastructure de cette innervation chez l’adulte réserve d’autres surprises.
L’étude en microscopie électronique et la reconstruction tri-dimensionnelle à partir de coupes sériées nous ont montré que l’innervation de la corne dorsale par les axones sérotoninergiques, dopaminergiques et noradrénergiques était en majorité non synaptique constituée de varicosités axonales remplies de vésicules, sans aucune structure post-synaptique. En revanche, l’innervation de la corne ventrale et celle de la colonne intermédiolatérale est conventionnellement synaptique [13]. La détection immunocytochimique des récepteurs correspondants a confirmé qu’ils étaient à distance des varicosités dans la corne dorsale, et en revanche contigus aux boutons axonaux dans la corne ventrale [14]. Ceci constitue l’exemple le mieux documenté à ce jour de ‘‘ transmission volumique ’’, qui réalise un mode paracrine d’interactions neuronales. Enfin, nous avons montré qu’un certain nombre de récepteurs sérotoninergiques de la corne dorsale étaient situés sur la surface d’astrocytes, suggérant que ces derniers interviennent dans des régulations à long terme de la transmission des messages sensitifs.
Nous avons ensuite exploré la plasticité de ces systèmes, en modifiant leurs cibles, par des lésions néonatales ou chez l’adulte. Les lésions des afférences sensitives entraînent une réorganisation spatiale des afférences sérotonergiques, dans les différentes couches de la corne dorsale de la moelle [15-16]. Par ailleurs, l’organisation non synaptique fait place à une projection majoritairement synaptique [17]. En revanche, les lésions des systèmes efférents de la colonne intermédiolatérale (projections sur la médullo-surrénale et le ganglion cervical supérieur) entraînent la disparition des afférences sérotoninergiques [18].
Cela illustre une différence fondamentale entre systèmes synaptiques, dont le maintien est lié à des facteurs trophiques originaires de la cible (NT3 dans le cas des neurones préganglionnaires) et des systèmes non-synaptiques, indépendants de ces cibles. Ces derniers constituent une variable ajustable d’une grande ampleur au sein du système nerveux central.
Un dernier volet de cette étude, plus physiologique, a consisté à étudier in vivo l’implication des systèmes monoaminergiques dans l’une des fonctions essentielles de la moelle épinière, la locomotion. Nous avons eu recours pour cela a une technique chirurgicalement délicate, mais d’une grande précision, la microdialyse chronique. Des micro-cannules implantées chroniquement pendant plusieurs semaines au niveau de la moelle thoraco-lombaire chez le rat permettent de mesurer par chromatographie liquide en temps réel la libération de neurotransmetteurs [19].
Nous avons pu ainsi démontrer que des rats non entravés courant sur des tapis roulant libéraient pendant l’exercice des quantités accrues de monoamines, avec une chronologie très précise par rapport à l’exercice, et ce non seulement au niveau de la corne antérieure [20] mais aussi de la corne dorsale, indiquant que la régulation de la locomotion par les monoamines implique également les afférences primaires sensitives.
Cette étude constitue la première preuve directe de libération in situ de monoamines en relation avec un exercice précis.
RÉPARATION MÉDULLAIRE
L’ensemble des travaux précédents nous a fourni les outils nécessaires à la mise en œuvre de stratégies de réparation médullaire. Ces stratégies se sont organisées autour de trois axes de recherche, qui constituent à terme une prise en charge raisonnée du traumatisé médullaire, et qui, par ailleurs, explorent un ensemble de paramètres de réparation du système nerveux susceptibles de concerner d’autres conditions pathologiques [21].
Le premier axe de recherche est la neuroprotection. L’étude des phénomènes post-lésionnels précoces a mis en évidence l’intervention de molécules toxiques responsable de l’aggravation progressive des lésions, et constituant la lésion dite secondaire post-traumatique ou post-accident vasculaire. Parmi ces toxiques, le glutamate joue un rôle essentiel, car il intervient très tôt et déclenche des cascades métaboliques conduisant à la libération d’autres toxiques, comme les radicaux libres.
En collaboration avec une équipe de chimistes, nous avons mis au point une série de molécules dérivées des phencyclidines qui présentent une affinité élevée pour l’un des récepteurs du glutamate, le récepteur NMDA [6]. Nous avons ensuite montré que ces molécules, qui sont des agonistes non-compétitifs sur ce récepteur, présentent des propriétés de neuro-protection sur des cultures neuronales primaires [22-23].
Une troisième étape de notre étude a consisté à étudier sur des modèles animaux lésionnels la neuroprotection apportée par une de ces molécules, la gacyclidine. Sur deux modèles, une lésion traumatique par lâcher de poids et une lésion vasculaire photochimique, toutes deux au niveau de la moelle thoracique basse, nous avons mis en évidence une neuroprotection significative, dont l’efficacité est maximale dans les deux heures suivant la lésion, et s’atténue ensuite [24-25].
Cette molécule a fait l’objet d’un essai clinique multicentrique en France, sur des blessés médullaires traités moins de deux heures après l’accident. Les résultats en cours de publication indiquent qu’un sous-groupe de patients, ceux présentant une lésion cervicale incomplète, bénéficie du traitement. D’autres essais cliniques sont programmés, en particulier, dans les traumatismes cérébraux.
Cette neuroprotection est essentielle, car elle conditionne les possibilités ultérieures de réparation, soit intrinsèque par repousse axonale, soit extrinsèque par thérapie substitutive cellulaire ou génique.
La régénération axonale est un objectif essentiel de la recherche contemporaine en neuroscience. Longtemps considérée comme impossible, après Cajal (1913), les travaux d’Aguayo et de ses collègues [5] ont montré que cette impossibilité n’était pas intrinsèque au neurone, mais bien plutôt due à un environnement non permissif au sein du système nerveux central lésé.
Nos travaux précédents sur le nerf optique nous avaient indiqué le rôle central de l’astrocyte dans la réaction tissulaire post-lésionnelle, et donc nous nous sommes tout naturellement tournés vers cette composante du milieu tissulaire. Nous avons étudié l’activation astrocytaire après une lésion médullaire, et montré qu’elle se caractérise par une augmentation rapide (moins d’une heure après la lésion) des ARNm de la protéine GFAP [26]. Cette augmentation des ARN est suivie en moins de 48 heures d’une augmentation massive de la synthèse de la protéine, dont la présence perdure après plusieurs semaines alors que les messagers ont retrouvé un taux normal.
Nous avons ensuite entrepris de bloquer localement, après une lésion, la réactivité astrocytaire au moyen d’un ester de cholestérol dont nous avions démontré qu’il était toxique in vitro pour les astrocytes tumoraux ou réactifs [27-28]. L’injection de cette molécule 48 heures après la lésion, à son voisinage immédiat, se traduit par un blocage de la réactivité astrocytaire, l’absence de formation d’une cicatrice impéné- trable, et la repousse de certains axones, en particulier sérotoninergiques [29].
Cette preuve de principe nous a amenés à approfondir cette problématique en nous dotant d’une série d’outils exclusifs, des souris transgéniques présentant l’inactivation des gènes codant pour les protéines constitutives de la cicatrice astrocytaire, vimentine et GFAP [30]. Après avoir étudié le phénotype de ces animaux [31-32] nous avons réalisé un modèle in vitro d’interaction neurone-astrocyte sous forme d’une co-culture d’astrocytes médullaires et de neurones embryonnaires corticaux, reproduisant une phénocopie de la situation d’une lésion médullaire. Nous avons montré de cette façon que les astrocytes dans lesquels le gène de la GFAP avait été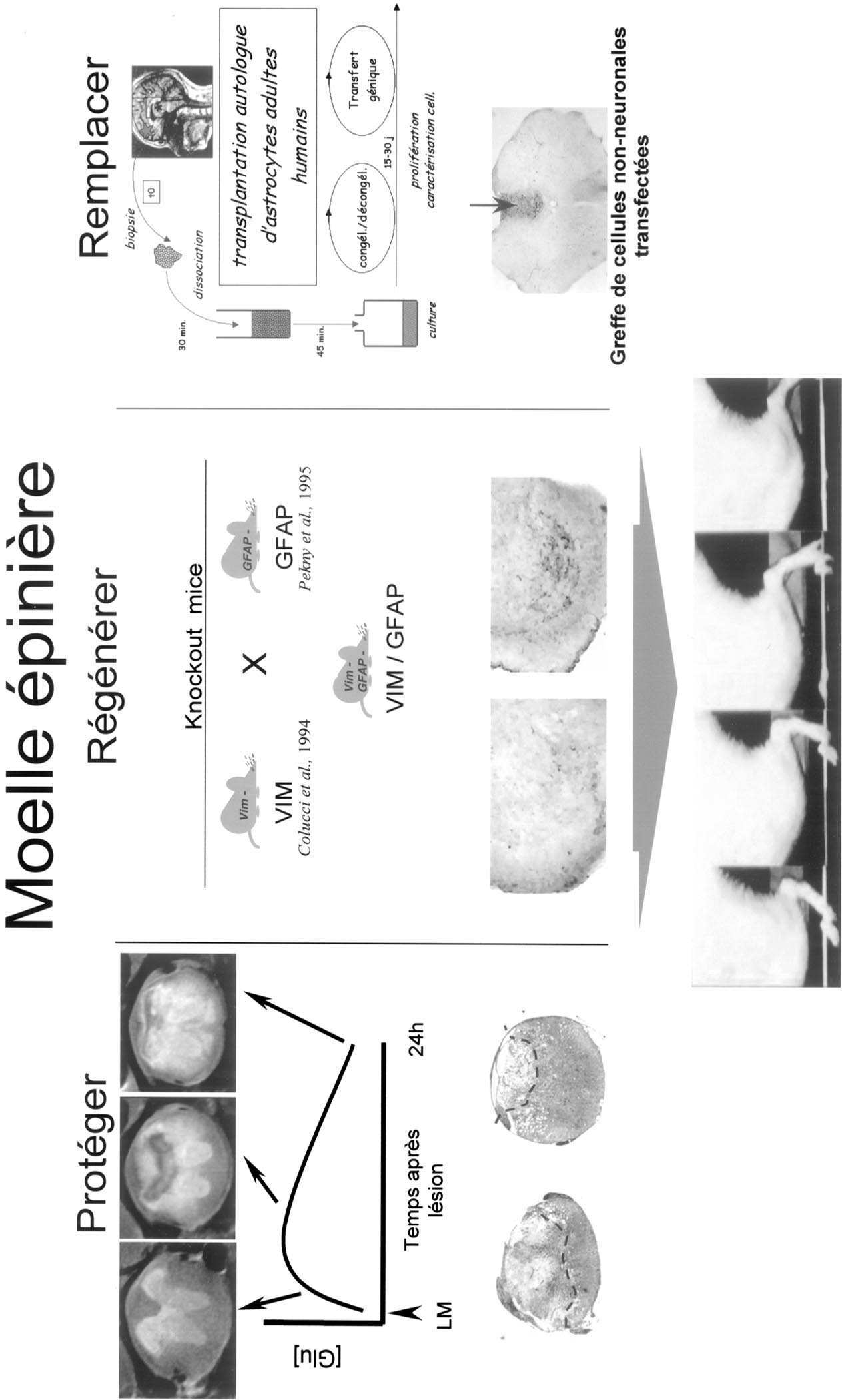
inactivé étaient particulièrement permissifs pour la survie des neurones et l’extension de leurs prolongements [33-34].
Nous avons étudié ces phénotypes in vivo sur un modèle d’hémisection médullaire, et nous avons montré que les animaux ayant subi une double inactivation des gènes codant pour la GFAP et la vimentine, présentent une cicatrice astrocytaire abortive, et une repousse axonale qui se traduit par une récupération significative des performances motrices dégradées par la lésion [35]. Nous étudions maintenant sur d’autres modèles lésionnels l’étendue de cette régénération et ses conséquences fonctionnelles. Par ailleurs, nous avons entrepris une étude de protéomique afin de mettre en évidence sur ce modèle l’ensemble des protéines dont la régulation est modifiée chez les mutants par rapport aux témoins. Cette étude doit pouvoir déboucher sur une stratégie thérapeutique précise fondée sur la thérapie génique ciblant les molécules essentielles pour cette repousse.
Le dernier volet de cette stratégie thérapeutique repose sur une approche de thérapie substitutive cellulaire et/ou génique. Nos travaux précédents nous avaient indiqué le rôle essentiel des monoamines, et en particulier de la sérotonine, dans la physiologie médullaire. Partant de ce constat, nous avons entrepris, dans un modèle de section médullaire complète, de transplanter, sous la lésion, des neurones embryonnaires susceptibles d’apporter à la moelle sous-lésionnelle les neurotransmetteurs absents.
La dissection fine des noyaux du raphé chez l’embryon fournit une population cellulaire qui se différencie in vitro en neurones sérotoninergiques [36]. Ces mêmes neurones embryonnaires injectés dans une moelle sous-lésionnelle, sept jours après la lésion, se développent au sein du tissu hôte et établissent progressivement des contacts synaptiques avec leurs cibles [37]. Ces contacts sont d’une grande spécificité, respectant la topologie des connexions établies entre les différents noyaux du raphé et les différentes cibles médullaires [38].
Sur le plan fonctionnel, nous avons constaté des modifications sensibles dans le comportement des animaux greffés. D’une part, les rats mâles ont récupéré un fonctionnement sexuel normal, avec la restauration d’une séquence érectionéjaculation directement sous le contrôle de l’innervation sérotonergique [39]. Par ailleurs, ces rats retrouvent une locomotion réflexe [40] dont l’origine se situe au niveau d’un centre locomoteur lombaire que nous avons pu localiser au niveau L2-L3, et dont nous apportons la première démonstration de sa fonctionnalité chez un mammifère adulte [41-42]. Cette étude ouvre la possibilité de rétablir des fonctions réflexes chez des paraplégiques — même anciens — au moyen de greffes cellulaires, similaires à celles utilisées chez les malades atteints de la maladie de Parkinson. L’utilisation de cette technique à une grande échelle nécessite de s’affranchir de l’utilisation de cellules embryonnaires humaines. Nous avons entrepris de mettre au point des stratégies substitutives fondées sur l’utilisation de cellules transfectées par génie génétique [43] ou de cellules pluripotentes précurseurs de cellules nerveuses [44].
CONCLUSION
Notre projet d’explorer la plasticité du système nerveux central des mammifères, et d’en utiliser les paramètres pour tenter de le réparer, est encore loin d’avoir atteint tous ses objectifs. Toutefois, nous avons pu dégager certains éléments prometteurs, susceptibles d’applications à court ou moyen terme dans une pathologie bien ciblée, celle des lésions traumatiques de la moelle épinière. Nous nous proposons, à partir de ce modèle, de démontrer le bien-fondé de notre approche pré-clinique, et d’étendre cette étude à d’autres pathologies aiguës ou chroniques du système nerveux.
REMERCIEMENTS
Ces travaux ont bénéficié de l’aide de l’IRME, de l’AFM, des associations Verticale et Demain Debout, et de la collaboration constante et efficace de M. Gimenez y Ribotta.
BIBLIOGRAPHIE [1] CAJAL S. R. — Estudios sobre la degeneracion y regeneracion del sistema nervioso. Madrid :
Moya (Ed.), 1913.
[2] RIO HORTEGA P.D. — Estudios sobre la neuroglia. Bol Real Soc Esp Hist Nat , 1921, 21 , 65-92.
[3] TERRAZAS P. — La neuroglia del cerebelo.
Rev Trimest Microgr , 1897, 1.
[4] CAJAL S. R. — Histologie du système nerveux de l’homme et des vertébrés. Paris : A. Maloine, 1909.
[5] AGUAYO A.J. — Axonal regeneration from injured neurons in the adult mammalian central nervous system. In : Synaptic plasticity, New York : C.W. Cotman Ed, 1985, 457-484.
[6] VIGNON J., PRIVAT A., CHAUDIEU I., THIERRY A., KAMENKA J. M., CHICHEPORTICHE R. — [3H] thienyl-phencyclidine ([3H]TCP) binds to two different sites in rat brain. Localization by autoradiographic and biochemical techniques. Brain Res , 1986, 378 , 133-41.
[7] RATABOUL P., FAUCON BIGUET N., VERNIER P., DE VITRY F., BOULARAND S., PRIVAT A., et al . —
Identification of a human glial fibrillary acidic protein cDNA : a tool for the molecular analysis of reactive gliosis in the mammalian central nervous system. J Neurosci Res , 1988, 20 , 165-75.
[8] RATABOUL P., VERNIER P., BIGUET N.F., MALLET J., POULAT P., PRIVAT A. — Modulation of GFAP mRNA levels following toxic lesions in the basal ganglia of the rat. Brain Res Bull , 1989, 22 , 155-61.
[9] PALKOVITS M., BROWNSTEIN M. J. — Microdissection of brain areas by the punch technique.
In : Cuello AC (Ed.), Brain microdissection techniques, Chichester, 1983, 1-36.
[10] RAJAOFETRA N., SANDILLON F., GEFFARD M., PRIVAT A. — Pre- and post-natal ontogeny of serotonergic projections to the rat spinal cord. J Neurosci Res , 1989, 22 , 305-21.
[11] RAJAOFETRA N., POULAT P., MARLIER L., GEFFARD M., PRIVAT A. — Pre- and postnatal development of noradrenergic projections to the rat spinal cord : an immunocytochemical study. Brain Res Dev Brain Res , 1992, 67 , 237-46.
[12] RAJAOFETRA N., RIDET J. L., POULAT P., MARLIER L., SANDILLON F., GEFFARD M., et al . —
Immunocytochemical mapping of noradrenergic projections to the rat spinal cord with an antiserum against noradrenaline. J Neurocytol , 1992, 21 , 481-94.
[13] RIDET J. L., RAJAOFETRA N., TEILHAC J. R., GEFFARD M., PRIVAT A. — Evidence for nonsynaptic serotoninergic and noradrenergic innervation of the rat dorsal horn and possible involvement of neuron-glia interactions. Neuroscience , 1993, 52 , 143-57.
[14] RIDET J.L., TAMIR H., PRIVAT A. — Direct immunocytochemical localization of 5-hydroxytryptamine receptors in the adult rat spinal cord : a light and electron microscopic study using an anti-idiotypic antiserum. J Neurosci Res , 1994, 38 , 109-21.
[15] MARLIER L., POULAT P., RAJAOFETRA N., PRIVAT A. — Modification of serotonergic immunoreactive pattern in the dorsal horn of the rat spinal cord following dorsal root rhizotomy.
Neurosci Lett , 1991, 128 , 9-12.
[16] MARLIER L., POULAT P., RAJAOFETRA N., PRIVAT A. — Modifications of serotonin-, substance P— and calcitonin gene-related peptide-like immunoreactivities in the dorsal horn of the spinal cord of arthritic rats : a quantitative immunocytochemical study. Exp Brain Res , 1991, 85 , 482-90.
[17] MARLIER L., POULAT P., RAJAOFETRA N., SANDILLON F., PRIVAT A. — Plasticity of the serotoninergic innervation of the dorsal horn of the rat spinal cord following neonatal capsaicin treatment. J Neurosci Res , 1992, 31 , 346-58.
[18] POULAT P., MARLIER L., PELLAS F., RAJAOFETRA N., PRIVAT A. — Effects of neonatal removal of superior cervical ganglion on serotonin and thyrotropin-releasing hormone immunoreactivity in the intermediolateral cell column of the rat spinal cord. Exp Brain Res , 1992, 91 , 21-8.
[19] GERIN C., LEGRAND A., PRIVAT A. — Study of 5-HT release with a chronically implanted microdialysis probe in the ventral horn of the spinal cord of unrestrained rats during exercise on a treadmill. J Neurosci Methods , 1994, 52 , 129-41.
[20] GERIN C., BECQUET D., PRIVAT A. — Direct evidence for the link between monoaminergic descending pathways and motor activity. I. A study with microdialysis probes implanted in the ventral funiculus of the spinal cord. Brain Res , 1995, 704 , 191-201.
[21] GIMÉNEZ Y RIBOTTA M., PRIVAT A. — Biological interventions for spinal cord injury.
Curr Opin
Neurol , 1998, 11 , 647-54.
[22] RONDOUIN G., DRIAN M.J., CHICHEPORTICHE R., KAMENKA J.M., PRIVAT A. — Noncompetitive antagonists of N-methyl-D-aspartate receptors protect cortical and hippocampal cell cultures against glutamate neurotoxicity. Neurosci Lett , 1988, 91 , 199-203.
[23] DRIAN M.J., KAMENKA J.M., PIRAT J.L., PRIVAT A. — Non-competitive antagonists of N-methyl-D-aspartate prevent spontaneous neuronal death in primary cultures of embryonic rat cortex. J Neurosci Res , 1991, 29 , 133-8.
[24] GAVIRIA M., PRIVAT A., d’ARBIGNY P., KAMENKA J., HATON H., OHANNA F. — Neuroprotective effects of a novel NMDA antagonist, Gacyclidine, after experimental contusive spinal cord injury in adult rats. Brain Res , 2000, 874 , 200-9.
[25] GAVIRIA M., PRIVAT A., d’ARBIGNY P., KAMENKA J. M., HATON H., OHANNA F. — Neuroprotective effects of gacyclidine after experimental photochemical spinal cord lesion in adult rats :
dose-window and time-window effects. J Neurotrauma , 2000, 17 , 19-30.
[26] MORIN-RICHAUD C., FELDBLUM S., PRIVAT A. — Astrocytes and oligodendrocytes reactions after a total section of the rat spinal cord. Brain Res , 1998, 783 , 85-101.
[27] BOCHELEN D., ECLANCHER F., KUPFERBERG A., PRIVAT A., MERSEL M. — 7 betahydroxycholesterol and 7 beta-hydroxycholesteryl-3-esters reduce the extent of reactive gliosis caused by an electrolytic lesion in rat brain. Neuroscience , 1992, 51 , 827-34.
[28] WERTHLE M., BOCHELEN D., ADAMCZYK M., KUPFERBERG A., POULET P., CHAMBRON J., et al .
— Local administration of 7 beta-hydroxycholesteryl-3-oleate inhibits growth of experimental rat C6 glioblastoma. Cancer Res , 1994, 54 , 998-1003.
[29] GIMÉNEZ y RIBOTTA M., RAJAOFETRA N., MORIN-RICHAUD C., ALONSO G., BOCHELEN D., SANDILLON F., et al . — Oxysterol (7 beta-hydroxycholesteryl-3-oleate) promotes serotonergic reinnervation in the lesioned rat spinal cord by reducing glial reaction.
J Neurosci Res , 1995, 41 , 79-95.
[30] GALOU M., COLUCCI-GUYON E., ENSERGUEIX D., RIDET J.L., GIMENEZ Y RIBOTTA M., PRIVAT A. et al . — Disrupted glial fibrillary acidic protein network in astrocytes from vimentin knockout mice.
J Cell Biol , 1996, 133 , 853-63.
[31] COLUCCI-GUYON E., GIMÉNEZ Y RIBOTTA M., MAURICE T., BABINET C., PRIVAT A. — Cerebellar defect and impaired motor coordination in mice lacking vimentin. Glia , 1999, 25 , 33-43.
[32] GIMÉNEZ Y RIBOTTA M., LANGA F., MENET V., PRIVAT A. — Comparative anatomy of the cerebellar cortex in mice lacking vimentin, GFAP, and both vimentin and GFAP. Glia , 2000, 31 , 69-83.
[33] MENET V., GIMÉNEZ Y RIBOTTA M., SANDILLON F., PRIVAT A. — GFAP null astrocytes are a favorable substrate for neuronal survival and neurite growth. Glia , 2000, 31 , 267-72.
[34] MENET V., GIMENEZ Y RIBOTTA M., CHAUVET N., DRIAN M.J., LANNOY J., COLUCCI-GUYON E., et al . — Inactivation of the glial fibrillary acidic protein gene, but not that of vimentin, improves neuronal survival and neurite growth by modifying adhesion molecule expression.
J Neurosci , 2001, 21 , 6147-58.
[35] GIMENEZ Y RIBOTTA M., MENET V., PRIVAT A. — The role of astrocytes in axonal regeneration in the mammalian CNS. Prog Brain Res , 2001, 132 , 587-610.
[36] PRIVAT A.P. — In vitro culture of serotonergic neurons from fetal rat brain.
J Histochem
Cytochem , 1982, 30 , 785-7.
[37] PRIVAT A., MANSOUR H., PAVY A., GEFFARD M., SANDILLON F. — Transplantation of dissociated fœtal serotonin neurons into the transected spinal cord of adult rats. Neurosci Lett , 1986, 66 , 61-6.
[38] RAJAOFETRA N., KONIG N., POULAT P., MARLIER L., SANDILLON F., DRIAN M. J., et al . — Fate of B1-B2 and B3 rhombencephalic cells transplanted into the transected spinal cord of adult rats : light and electron microscopic studies. Exp Neurol , 1992, 117 , 59-70.
[39] PRIVAT A., MANSOUR H., GEFFARD M. — Transplantation of fetal serotonin neurons into the transected spinal cord of adult rats : morphological development and functional influence. Prog Brain Res , 1988, 78 , 155-66.
[40] YAKOVLEFF A., ROBY-BRAMI A., GUEZARD B., MANSOUR H., BUSSEL B., PRIVAT A. — Locomotion in rats transplanted with noradrenergic neurons. Brain Res Bull , 1989, 22 , 115-21.
[41] FERABOLI-LOHNHERR D., ORSAL D., YAKOVLEFF A., GIMENEZ Y RIBOTTA M., PRIVAT A. — Recovery of locomotor activity in the adult chronic spinal rat after sublesional transplantation of embryonic nervous cells : specific role of serotonergic neurons. Exp Brain Res , 1997, 113 , 443-54.
[42] GIMÉNEZ Y RIBOTTA M., PROVENCHER J., FERABOLI-LOHNHERR D., ROSSIGNOL S., PRIVAT A., ORSAL D. — Activation of locomotion in adult chronic spinal rats is achieved by transplantation of embryonic raphe cells reinnervating a precise lumbar level. J Neurosci , 2000, 20 , 5144-52.
[43] LEVALLOIS C., PRIVAT A., MALLET J. — Adenovirus insertion encoding the Lac Z gene in human nervous cells in primary dissociated cultures. C R Acad Sci III , 1994, 317 , 495-8.
[44] GUILLEMAIN I., ALONSO G., PATEY G., PRIVAT A., CHAUDIEU I. — Human NT2 neurons express a large variety of neurotransmission phenotypes in vitro . J Comp Neurol , 2000, 422 , 380-95.
DISCUSSION
M. Raymond HOUDART
Qu’en est-il des recherches initiées il y a une dizaine d’années par l’Institut de recherche sur la moelle épinière (IRME) ?
Les travaux que je viens de vous exposer sont la suite directe de ceux qui ont été initiés par l’IRME, il y a plus de dix ans. Depuis nous avons bénéficié constamment de l’aide de cette institution, qui a permis de rassembler les équipes françaises travaillant dans ce domaine, et de développer leur complémentarité.
M. Roger NORDMANN
Vous avez cité le GK 11 en tant qu’antagoniste non compétitif du NMDA. Cette action est-elle localisée à la moelle épinière ou est-elle également présente au niveau cérébral ?
L’alcoolisation des rongeurs a pour conséquence des altérations du glutamate et des récepteurs NMDA (variables selon qu’il s’agit d’alcoolisation aiguë ou chronique), dont le rôle dans l’appétence envers l’alcool, les phénomènes de dépendance ainsi que les destructions neuronales reste encore controversé. Ne pensez-vous pas que l’utilisation du GK 11 permettrait de préciser ce rôle ?
Le GK11 se comporte comme un antagoniste du NMDA non seulement au niveau de la moelle, mais aussi de l’ensemble du système nerveux central. Je retiens la suggestion qui nous est faite de l’utiliser dans la lutte contre les dépendances, l’alcool en particulier.
Certains travaux sur des molécules de la famille des phencyclidines à laquelle appartient le GK11, ont montré des effets intéressants dans le traitement de la dépendance à la cocaïne.
M. Jean-Jacques HAUW
Comme vous l’avez confirmé, la régénération axonale dépend de cellules névrogliales et notamment des astrocytes. Un certain nombre d’arguments laisse penser que les astrocytes ont des propriétés différentes selon leur topographie. Pensez-vous que ces différences pourraient être utilisées pour ouvrir de nouvelles pistes thérapeutiques ?
Nous avons constaté des différences entre les astrocytes provenant de diverses régions du système nerveux. Dans des expériences de co-culture neurones / astrocytes, les astrocytes corticaux se comportent un peu différemment des astrocytes médullaires, ces derniers apparaissant comme les plus obstructifs à une régénération. Nous entreprenons actuellement une analyse protéomique de divers types d’astrocytes et de cellules apparentées, dans le but d’identifier des cibles thérapeutiques originales dans le domaine de la régénération.
M. Jean-Marie BOURRE
Pour ce qui concerne la greffe cellulaire, le clonage « thérapeutique » étant, semble-t-il, exclu pour des raisons de mise en œuvre trop longue, vers quelles cellules souche l’avenir est-il ? Dans quels délais peut-on espérer une application clinique de la greffe de cellule ?
Dans cette problématique de greffe ‘‘ substitutive ’’, nous nous orientons vers des cellules souches pluripotentes, issues de tissus adultes, en particulier le muscle et la moelle osseuse, qui, après expansion en culture, sont susceptibles, pour certaines d’entre elles de se différencier en neurones. Nous pourrions ainsi réaliser des autogreffes chez les blessés médullaires. Mais nous sommes loin de maîtriser totalement la différenciation de ces cellules. Il est toujours difficile de fixer un délai. Nous savons chez l’animal que des cellules greffées plusieurs semaines après l’accident rétablissent une fonction. L’extrapolation à l’homme donnerait un délai de plusieurs mois, voire d’années, mais ce n’est qu’une extrapolation. Pour ce qui est de la mise en œuvre d’un essai clinique pilote, qui utiliserait des cellules embryonnaires humaines, nous y travaillons avec les comités d’éthique.
M. Alain LARCAN
À côté de l’intérêt majeur de la neuroprotection dans les traumatisme médullaires, on sait que la même substance (GK11) est efficace dans les intoxications par organophosphores, ce qu’ont démontré les chercheurs du CRESSA à Grenoble. Peut-on distinguer un seuil d’irréversibilité des lésions permettant de prévoir l’échec des traitements préventifs et curatifs ? Pouvez-vous préciser l’orientation de la multithérapie, nouveaux antagonistes des A.A excitateurs, inhibiteurs calciques, antiradicaux libres ?
Nous travaillons depuis de nombreuses années avec nos collègues de Grenoble, qui ont effectivement démontré, de façon très convaincante, sur des petits mammifères mais aussi des primates, que le GK11 pourrait être un protecteur efficace contre les gaz de combat organophosphorés (SARIN, SOMAN). Leurs travaux indiquent qu’une intervention dans les deux heures qui suivent l’intoxication en réduit considérablement la gravité, et en particulier les séquelles cérébrales. La multithérapie associera le GK11 ou un analogue avec un piégeur de radicaux libres, et peut être un antagoniste AMPA, mais ceux dont nous disposons passent mal la barrière hémato-encéphalique.
M. Claude DREUX
En cas de section complète de la moelle, le transfert de noyaux du raphé hétérologue pourrait-il être envisagé pour le repeuplement des cibles normales ?
Dans le cas en particulier de la maladie de Parkinson et de la chorée de Huntington, l’hétérogreffe implique un traitement immunosuppresseur permanent, dont l’indication ne se discute pas s’agissant de maladies évolutives, dont le pronostic est parfois gravissime. Il n’en est pas de même pour les blessés médullaires. C’est pour cette raison, entre autres, que nous nous orientons à terme vers une thérapie autologue qui ferait moins courir de risques d’effets secondaires aux patients.
M. Pierre RONDOT
Pourquoi avoir greffé essentiellement des neurones sérotoninergiques dans le segment terminal d’une moelle sectionnée ?
Le choix des neurones sérotoninergiques est fondé sur une série de travaux de notre laboratoire et des groupes suédois et danois (Grillner, Hultborn) qui ont montré que la sérotonine jouait un rôle essentiel dans le contrôle de l’activité du motoneurone responsable de la locomotion, au moins chez les petits rongeurs. Il semble d’ailleurs en être de même chez les primates.
M. Georges SERRATRICE
Qu’en est-il de la réorganisation des « patrons spinaux » ?
Notre approche de greffe est justement fondée sur la réactivation des ‘‘ patrons spinaux ’’, dont l’existence, postulée depuis longtemps, a été confirmée chez le rat nouveauné, par le groupe de F. Clarac à Marseille et chez l’adulte par nous-mêmes. De façon très intéressante, ces ‘‘ patrons spinaux ’’, existant avant la naissance chez le rat mais aussi chez l’homme, paraissent extrêmement robustes. Les travaux électrophysiologiques de Dimitrijevic ont montré qu’ils étaient toujours activables chez des paraplégiques anciens.
Syndrome de Turner et mosaïcisme
Turner’s syndrome and mosaicism
Jacques BATTIN *
* Membre correspondant de l’Académie nationale de médecine .
Service de Génétique Médicale — Hôpital des Enfants — CHU Pellegrin — Place Amélie Raba-Léon — 33076 Bordeaux cedex.
Tirés-à-part : Professeur Jacques BATTIN, à l’adresse ci-dessus.
Article reçu le 14 octobre 2002, accepté le 13 janvier 2003.
* INSERM U336, E.P.H.E. Université de Montpellier II, Place E. Bataillon — cc 106 — 34095 Montpellier cedex 5. Tirés-à-part : Professeur Alain PRIVAT, à l’adresse ci-dessus. Article reçu le 6 août 2002, accepté le 2 décembre 2002.
Bull. Acad. Natle Méd., 2003, 187, n° 2, 345-357, séance du 4 février 2003