
Résumé
La listériose est une infection sévère atteignant l’homme et la plupart des espèces animales, causée par une bactérie à Gram positif, Listeria monocytogenes. Ce pathogène à croissance intracellulaire facultative est largement répandue dans la nature et infecte l’homme par la nourriture contaminée. Après ingestion, les bactéries peuvent franchir la muqueuse digestive et disséminer par voie sanguine. Les bactéries peuvent alors envahir le système nerveux central et le placenta, causant des méningo-encéphalites et des infections fœto-placentaires sévères. Ce pouvoir invasif de L. monocytogenes explique la forte mortalité de la maladie estimée à environ 30 % malgré l’antibiothérapie. La listériose constitue un modèle remarquable pour étudier comment une bactérie pathogène peut traverser les barrières hémoencéphalique et placentaire. Les mécanismes moléculaires du parasitisme intracellulaire de L. monocytogenes et de sa diffusion dans les tissus de l’hôte sont aujourd’hui les mieux compris parmi les bactéries pathogènes.
Summary
Listeriosis is a severe infectious disease occurring both in humans and in most other animal species. It is caused by a Gram-positive bacterium, Listeria monocytogenes. This facultative intracellular pathogen is widespread in nature and infects humans through contaminated food. After ingestion, bacteria can cross the gut mucosa and disseminate via the bloodstream. They may then invade the central nervous system and the placenta, causing severe meningoencephalitis and fetoplacental infections, respectively. The mortality rate is about 30 %, despite antibiotic therapy. Listeriosis offers an excellent model for studying how bacterial pathogens can cross the blood-brain barrier and placenta. Our understanding of the molecular mechanisms of intracellular parasitism and dissemination to host tissues is better for L. monocytogenes than for any other bacterial pathogen.
INTRODUCTION
Listeria monocytogenes est un des pathogènes dont les mécanismes moléculaires de la pathogénicité ont été bien éclaircis ces dernières années. C’est une bactérie à Gram positif qui est responsable de la listériose, une maladie sporadique pouvant atteindre la plupart des espèces animales, incluant l’homme et les mammifères, les oiseaux et même les animaux à sang froid [1]. Ce pouvoir pathogène ubiquiste témoigne de mécanismes de virulence relativement universels. La gravité de la listériose est liée à la capacité des bactéries de pénétrer le système nerveux central et le placenta [2, 3].
La mortalité globale de la maladie est estimée à 25-30 % avec des séquelles neurologiques très fréquentes (40 %) en dépit de l’antibiothérapie [4]. Après Escherichia coli K1 et Streptococcus agalactiae , L. monocytogenes est une cause majeure d’infections néonatales, responsable de près de 1 % des méningites bactériennes et de 0,15 % de la mortalité périnatale.
Incidence et épidémiologie de la listériose
La listériose est une maladie relativement rare, plus fréquente chez les femmes enceintes qui représentent plus de la moitié des cas de listériose et les patients aux défenses immunitaires affaiblies. La maladie est souvent insidieuse avec fièvre, frissons, céphalées, parfois pharyngites ou syndrome pseudo-grippal, précédant les signes de méningite et de rhombencéphalite avec raideur méningée et atteinte des nerfs crâniens [5]. Cette méningo-encéphalite très fréquente chez le nouveau-né survient aussi dans près de deux tiers des cas chez le grand enfant et l’adulte.
L’incidence de la maladie surtout observée dans les pays industrialisés oscille entre 2-15 cas/ million d’habitants [4]. La bactérie très répandue dans la nature (sol, eaux usées, ensilages…) où elle vit à l’état saprophyte [1] contamine l’homme et les animaux par ingestion d’aliments souillés où les bactéries ont proliféré. Le renforcement des mesures d’hygiène et de prévention par les professionnels de l’industrie agro-alimentaire a permis de diminuer l’incidence de la listériose dans les pays industrialisés. En France où la listériose est à déclaration obligatoire depuis 1998, l’incidence annuelle est estimée à environ 200 cas diagnostiqués. Dans les rares pays comme la France, la Grande-Bretagne ou les États-Unis, où existent des systèmes efficaces de surveillance épidémiologique, on observe des épidémies habituellement limitées, survenant à la suite de contaminations accidentelles d’un aliment particulier souvent distribué sur l’ensemble du territoire. Les épidémies sont habituellement détectées par le centre de référence de l’Institut Pasteur par la mise en évidence de plusieurs cas liés à des isolats bactériens génétiquement identiques. En France depuis 1992, on a dénombré sept épidémies dues à des charcuteries (rillettes, langue de porc…) et des fromages à patte molle ou au lait cru. Dans le monde, on a aussi identifié des produits de la mer et des produits végétaux à l’origine d’épidémies [2, 4].
On a récemment aussi décrit une nouvelle forme clinique de listériose jusqu’ici
méconnue. Contrairement à la listériose classique des sujets fragiles qui n’entraîne pas de diarrhée, c’est une toxi-infection alimentaire avec diarrhée et fièvre survenant chez des adultes jeunes immunocompétents quelques heures après ingestion de salades de maïs ou de soja massivement contaminés [6-8].
Le parasitisme intracellulaire de L. monocytogenes
Une des caractéristiques remarquables de
L. monocytogenes est sa capacité de survivre et de se multiplier dans la plupart des cellules de l’organisme, incluant les macrophages [9], à l’exception des polynucléaires qui la détruisent rapidement. Aux différentes phases du processus infectieux in vivo , les cellules parasitées constituent des sanctuaires où les microorganismes sont à l’abri du système immunitaire [10]. Ce parasitisme intracellulaire est lié à la production de facteurs de virulence qui sont des protéines exposées à la surface des bactéries ou sécrétées dans leur microenvironnement. Nous avons contribué à la compréhension de ces mécanismes en mettant en évidence plusieurs de ces facteurs de virulence, notamment la listériolysine O, ActA et l’internaline. Dans une première étape, les bactéries entrent en contact étroit avec les cellules grâce à des protéines attachées à la paroi bactérienne, les internalines A et B [11], qui interagissent avec des récepteurs cellulaires tels que la E-cadhérine [12], induisant un processus de phagocytose. À l’intérieur des cellules, les bactéries se trouvent dans des phagosomes acidifiés où elles sont exposées aux réactions microbicides [9]. Elles peuvent échapper des vacuoles en détruisant les membranes phagosomales grâce à une exotoxine hémolytique, la listériolysine O [13-15] et à des phospholipases C [16]. Les bactéries accèdent ainsi au cytoplasme où elles se multiplient et polymérisent l’actine, un constituant abondant du cytosquelette. Cette polymérisation est due à la présence d’une autre protéine bactérienne de surface, ActA [17], qui reconnaît spécifiquement certaines protéines cellulaires (profiline, vinculine…) jouant le rôle d’intermédiaires dans le processus de « nucléation » de l’actine. Ceci propulse les bactéries qui forment des comètes d’actine par protrusion vers les cellules adjacentes où elles sont entourées d’une double membrane cytoplasmique. Les microorganismes vont échapper de cette nouvelle vacuole en produisant une phospholipase [18]. Ce mécanisme de dissémination permet de prolonger la survie intracellulaire de L monocytogenes à l’abri du système immunitaire (Figure 1 A d’après [24]). La survie intracellulaire de L. monocytogenes résulte donc d’un phénomène de compétition entre la rapidité de mise en œuvre de la fusion phagolysosomale et la capacité des bactéries d’échapper rapidement au phagosome [9]. Les protéines de stress jouent aussi un rôle majeur dans la survie et l’échappement des phagosomes [19-23]. L’ensemble des gènes codant pour les facteurs de virulence est réparti sur un petit fragment du chromosome bactérien en un îlot de pathogénicité contrôlé par l’activateur transcriptionnel PrfA (Figure 1 B).
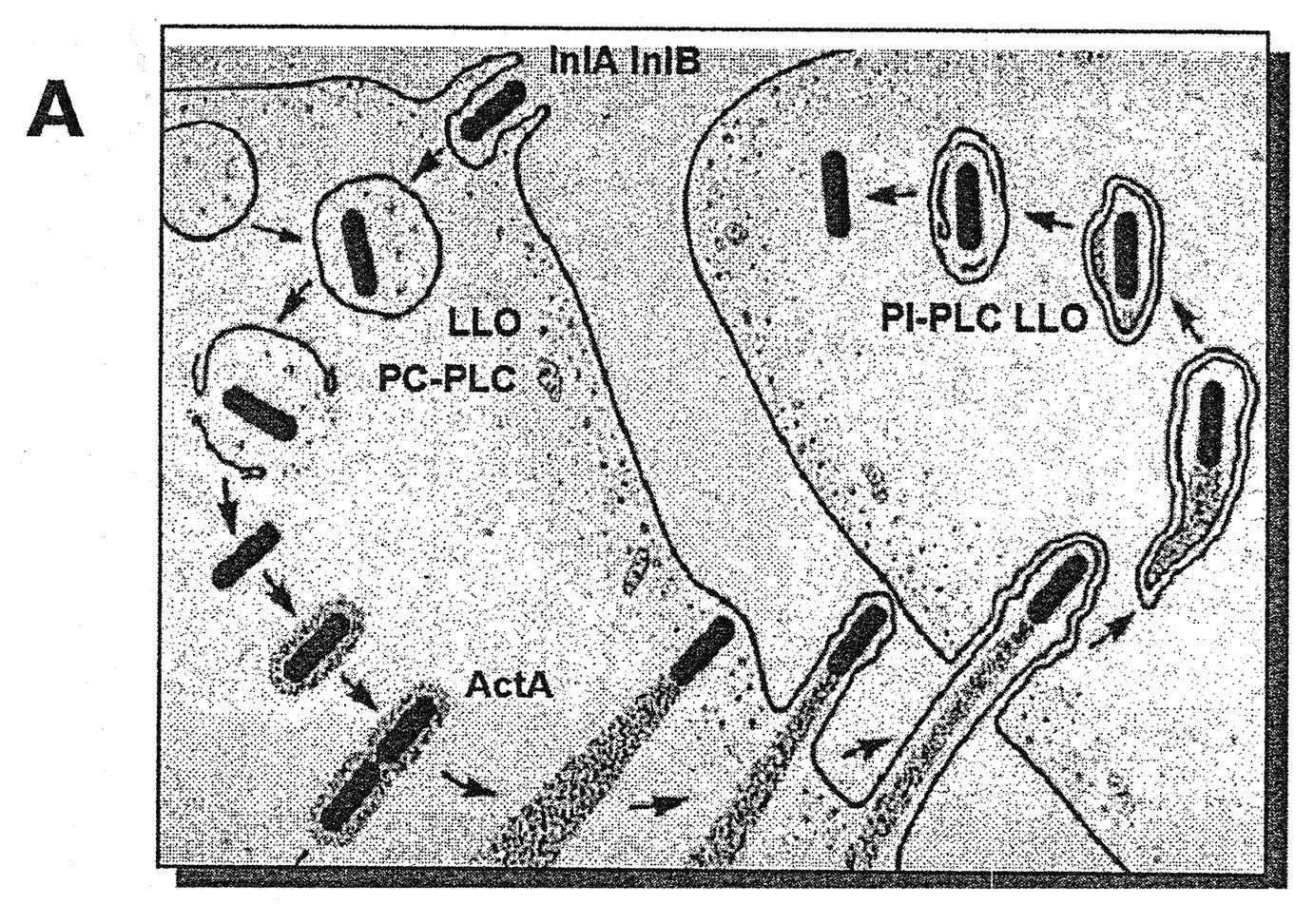
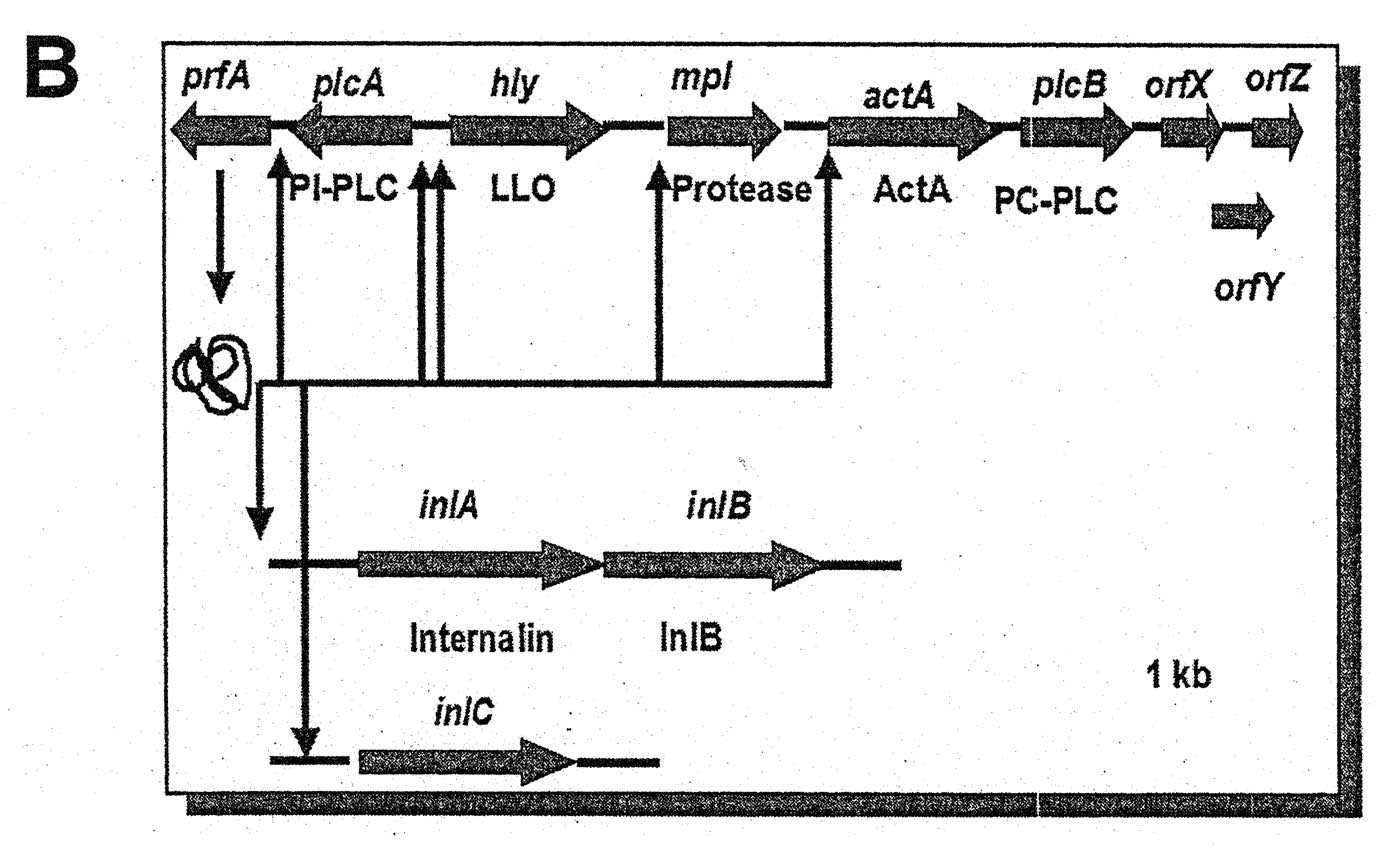
FIG. 1 A — Cycle de réplication intracellulaire de
Listeria monocytogenes , d’après [24].
FIG. 1 B. — L’ilôt de pathogénicité rassemblant les gènes de virulence de
Listeria monocytogenes sous contrôle de l’activateur PrfA.
Physiopathologie de la listériose humaine
L’invasion par voie digestive
À partir des données anatomopathologiques et de modèles expérimentaux murins, on a pu reconstituer le scénario de la listériose (Figure 2). Après ingestion d’aliments contaminés, les bactéries peuvent franchir la muqueuse digestive en s’infiltrant au niveau des entérocytes et des plaques de Peyer, où elles sont ingérées par des cellules dendritiques qui les véhiculent vers la circulation lymphatique puis sanguine [25, 26]. Les bactéries circulantes sont captées par les macrophages résidents du foie, de la rate et aussi, comme nous l’avons très récemment montré, de la moelle osseuse qui est infectée dès les premières heures [27]. Nous avons découvert que la moelle osseuse constitue un important réservoir infectieux, où les bactéries se multiplient rapidement dans les cellules de la lignée myélo-monocytaire [27].
Dans les capillaires sinusoïdes du foie, les bactéries présentes dans la circulation pénètrent dans les hépatocytes directement ou par l’intermédiare des cellules de Küpffer infectées. L’internaline joue un rôle crucial dans ce processus [28]. Les bactéries se multiplient dans les hépatocytes et disséminent rapidement de proche en proche. Ceci entraîne une réponse inflammatoire intense, avec formation de granulomes constitués de polynucléaires, de monocytes et de lymphocytes dès les premiè- res heures de l’infection. L’immunité protectrice est de nature cellulaire et implique le recrutement de lymphocytes CD-8 cytotoxiques. La maladie à ce stade est souvent contrôlée chez les sujets immunocompétents qui feront une infection a minima, souvent asymptomatique. Ceci est probablement le scénario le plus fréquent, si l’on considère la fréquence de l’exposition à
L. monocytogenes et la rareté de la maladie clinique. Cependant, si l’inoculum a été massif ou chez les femmes enceintes et les patients immunodéprimés (sida, patients sous chimiothérapie, patients neutropéniques), ou présentant des anomalies hépatiques (cirrhose, hémachromatose), l’infection n’est pas contrôlée et une bactériémie survient, exposant le système nerveux central et le placenta.
L’infection du système nerveux central
La listériose neuro-méningée se présente chez l’homme sous la forme d’une méningite lympho-monocytaire ou purulente. La localisation méningée est associée dans près de la moitié des cas à des signes d’encéphalite diffuse avec des abcès intracéré- braux localisés de façon prédominante au rhombencéphale. Des abcès nécrotiques coalescents et multiples apparaissent entraînant des pertes de substances importantes du tissu nerveux. Plus rarement, il peut s’agir de formes encéphalitiques pures sans méningite. Chez le nouveau-né, l’atteinte méningée prédomine.
Tout porte à croire que le cerveau est infecté par voie hématogène par passage des bactéries à travers les endothéliums des capillaires cérébraux. On sait que la bacté- riémie est requise pour l’envahissement cerébral [29], les bactéries étant surtout associées aux monocytes circulants [30, 31]. La bactériémie s’accompagne d’une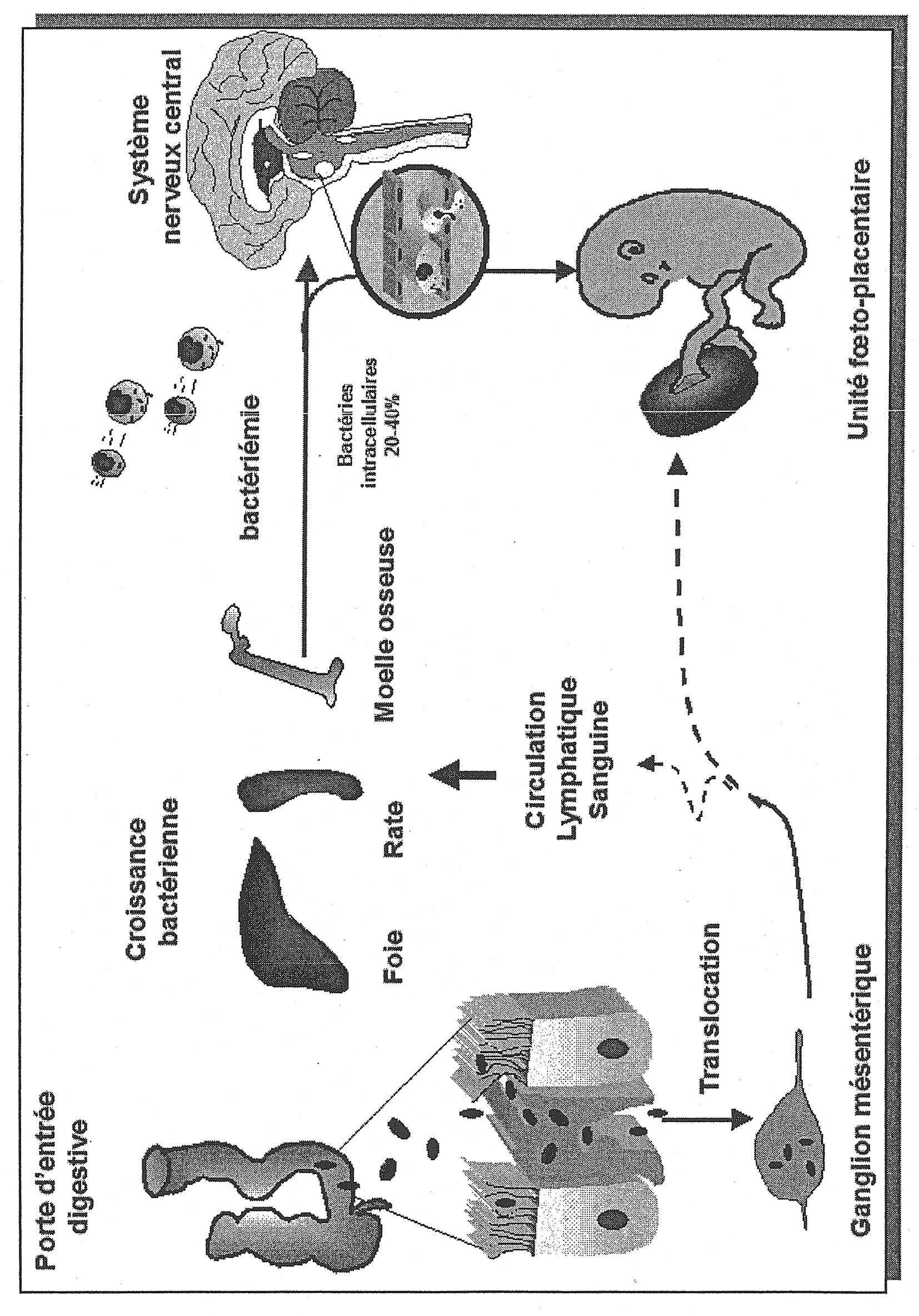 .
.
é riose list la de gique tholo ysiopa ph é nario Sc — 2.
IG.
F
forte activation des endothéliums cérébraux liée à la production de listériolysine O [32, 33]. Ceci est dû à l’activation par cette exotoxine du facteur transcriptionnel cellulaire NF-kB contrôlant de nombreux gènes de l’inflammation. Il en résulte une expression par les cellules endothéliales de molécules d’adhésion (ICAM-1 et E-sélectine) et de chimiokines à puissant effet chémo-attractant pour les cellules inflammatoires (IL-8 et MCP-1) [32, 33]. Ainsi, les cellules inflammatoires circulantes sont recrutées et adhèrent fortement aux endothéliums des capillaires cérébraux [27]. Nous avons pu démontrer très récemment que les cellules myélo-monocytaires infectées de la moelle osseuse sont mobilisés dans le sang et gagnent les capillaires cérébraux qu’elles franchissent par diapédèse [27]. La barrière hémo-encéphalique est donc franchie par des monocytes médullaires véhiculant des bactéries intracellulaires vivantes, en jouant le rôle de « cheval de Troie ».
L’infection materno-infantile
La contamination de la mère se fait par voie orale. La gestation sensibilise à l’infection en diminuant les défenses immunitaires, surtout au cours du 3ème trimestre de la grossesse. Les signes cliniques chez la mère sont souvent inapparents, ou résumés à un syndrome pseudogrippal avec fièvre et frissons, fatigue, maux de tête et myalgies qui peuvent précéder l’accouchement de 2 à 14 jours ou plus. Une rechute fébrile avec bactériémie est souvent observée au cours de l’accouchement. La plupart des cas de listériose sont décrits après le 5ème mois de grossesse avec accouchement prématuré, mais des avortements spontanés ou répétés peuvent survenir avant cette date.
L. monocytogenes peut infecter l’enfant le plus souvent par voie placentaire, à la suite d’une bactériémie survenant chez la mère [34]. L’infection in utero est caracté- risée par une colonisation du placenta où de multiples abcès granulomateux se forment dans les villosités trophoblastiques et la decidua . Plus tardivement au cours de la grossesse, des foyers granulomateux sont visibles à la surface du placenta. Cette infection est rapidement associée à une chorioamniotite et à une infection sévère de l’enfant in utero évidente dès la naissance avec cyanose, apnée, détresse respiratoire et coma. Une pneumonie péri-bronchiale ou associée à des infiltrats diffus, plus rarement miliaires, est souvent retrouvée. Dans certaines formes très sévères ( granulomatosis infantiseptica ), on observe une granulomatose disséminée à la plupart des organes, tout particulièrement le foie, les poumons et les méningites, mais aussi la rate, le tube digestif et l’œsophage, les organes lymphoïdes, les surrénales, les muscles (squelettiques, myocarde) et la peau. Dans ces formes graves, évoluant depuis plusieurs jours, la mortalité est élevée (parfois 50 à 75 %). Dans un faible nombre de cas (< 10 %), le nouveau né est contaminé juste avant ou au cours de l’accouchement sans qu’il y ait d’infection placentaire, par ingestion du liquide amniotique infecté ou au cours du passage de la filière génitale (< 10 % des cas). Il présente alors une infection différée de 8 à 60 jours après la naissance, d’assez bon pronostic car précocement diagnostiquée. Le rôle des différents facteurs de virulence dans l’envahissement du placenta est en cours d’étude.
Conclusion
Dans la nature, il existe au moins deux types de pathogènes. Certains se sont progressivement adaptés parfois de façon très spécifique à certaines espèces vivantes au cours de milliers d’années d’évolution, tels que le vibrion cholérique, le bacille typhique, le bacille de la lèpre et celui de la tuberculose pour l’espèce humaine.
D’autres infectent accidentellement l’homme ou les animaux, par exemple les bactéries du sol donnant le tétanos ou le botulisme. L. monocytogenes appartient à cette seconde catégorie. Infectant l’homme accidentellement par les aliments contaminés, cette bactérie est mal adaptée au tube digestif nécessitant des doses infectantes massives pour induire l’infection clinique. En revanche, les quelques bactéries qui franchissent la barrière digestive sont très virulente du fait de leur aptitude à parasiter les cellules des hôtes infectés. L’îlot de pathogénicité rassemblant les gènes responsables du parasitisme intracellulaire a été sélectionné au cours de l’évolution pour permettre à cette bactérie saprophyte de survivre dans l’environnement, notamment dans les amibes prédatrices qui pullulent dans les eaux usées et le sol.
C’est par hasard que ces mêmes gènes sont utilisés efficacement pour parasiter les macrophages et les cellules des animaux infectés et pour favoriser la dissémination des bactéries en les véhiculant dans les cellules monocytaires sanguines vers le placenta et le système nerveux central.
BIBLIOGRAPHIE [1] GRAY M. L., KILLINGER A. H. — Listeria monocytogenes and listeric infections.
Bacteriol. Rev , 1966, 30 , 309-382.
[2] VASQUEZ-BOLAND J.A., KUHN M., BERCHE P., CHAKRABORTY T., DOMINGUEZ-BERNAL G., GOEBEL W., GONZALEZ-ZORN B., WEHLAND J., KREFT J. — Listeria pathogenesis and molecular determinants. Clin. Microbiol. Rev. , 2001, 14 , 584-640.
[3] ROUQUETTE C., BERCHE P. — Pathogenesis of infection by Listeria monocytogenes.
Int. Micro- biol. , 1996, 12 , 245-258.
[4] FARBER J.M., PETERKIN P.I. — Listeria monocytogenes, a food-borne pathogen.
Microbiol.
Rev. , 1991, 55 , 476-511.
[5] LAVETTER A., LEEDOM J.M., MATHIES A.W., IVLER D., WEHRLE P.F. — Meningitis due to Listeria monocytogenes. A review of 25 cases. N. Engl. J. Med. , 1971, 285 , 598-603.
[6] SALAMINA G., DALLE DONNE E., NICCOLINI A., PODA G., CESARONI D., BUCCI M., FINI R., MALDINI M., SCHUCHAT A., SWAMINATHAN B., BIBB W., ROCOURT J., BINKIN N., SALMASO S.A.
— Foodborne outbreak of gastroenteritis involving Listeria monocytogenes. Epidemiol. Infect. , 1996, 117, 429-436.
[7] AURELI P., GIOVANNI C.F., CAROLI D., MARCHIARO G., NOVARA O., LEONE L., and SALMASO S.
— An outbreak of febrile gastroenteritis associated with corn contaminated by Listera monocytogenes . N. Engl. J. Med. , 2000, 342, 1236-1241.
[8] DALTON C.B., AUSTIN C.C., SOBEL J., HAYES P.S., BIBB W.F., GRAVES L.M., SWAMINATHAN B., PROCTOR M.E., GRIFFIN P.M. — An outbreak of gastroenteritis and fever due to Listeria monocytogenes in milk . N. Engl. J. Med., 1997, 336 , 100-105.
[9] DE CHASTELLIER C., BERCHE P. — Fate of Listeria monocytogenes in murine macrophages :
evidence for simultaneous killing and survival of intracellular bacteria. Infect. Immun., 1994, 62 , 543-553.
[10] GAILLARD J.L., BERCHE P., FREHEL C., GOUIN E., COSSART P. — Entry of Listeria monocytogenes into cells is mediated by a repeat protein analogous to surface antigens from Grampositive extracellular pathogen. Cell , 1991, 65 , 1127-1141.
[11] MENGAUD J., OHAYON P., GOUNON R., MÈGE M., COSSART P. — E-cadherin is the receptor for internalin, a surface protein required for entry of Listeria monocytogenes into epithelial cells.
Cell. , 1996, 84 , 923-932.
[12] GAILLARD J.L., BERCHE P., SANSONETTI P. — Transposon mutagenesis as a tool to study the role of hemolysin in the virulence of Listeria monocytogenes. Infect. Immun ., 1986, 52 , 50-55.
[13] GAILLARD J.L., BERCHE P., MOUNIER J., RICHARD S., SANSONETTI P. —
In vitro model of penetration and intracellular growth of Listeria monocytogenes in the human enterocyte-like cell line Caco-2. Infect. Immun. , 1987 , 55 , 2822-2829.
[14] GEOFFROY C., GAILLARD J.L., ALOUF J.E., BERCHE P. — Purification, characterization and toxicity of sulfhydryl-activated hemolysin (listeriolysin O) from Listeria monocytogenes. Infect.
Immun., 1987, 55 , 1641-1646.
[15] COSSART P., VICENTE M.F., MENGAUD J., BAQUERO F., PEREZ-DIAZ J.C., BERCHE P. — Listeriolysin O is essential for virulence of Listeria monocytogenes : direct evidence obtained by gene complementation. Infect. Immun., 1989 , 57 , 3629-3636.
[16] GEOFFROY C., RAVENEAU J., BERETTI J.L., LECROISEY A., ALOUF J.E., BERCHE P. — Purification and characterization of an extracellular 29-kDa phosphatidylcholine-phospholipase C from Listeria monocytogenes. Infect. Immun., 1991, 59 , 2382-2388.
[17] KOCKS C., GOUIN E., TABOURET M., BERCHE P., OHAYON H., COSSART P. — Listeria monocytogenes induced actin assembly requires the act A gene product, a surface protein. Cell. , 1992, 68 , 521-531.
[18] VAZQUEZ-BOLAND J.A., KOCKS C., DRAMSI S., OHAYON H., GEOFFROY C., MENGAUD J., COSSART P. — Nucleotide sequence of the lecithinase operon of Listeria monocytogenes and possible role of lecithinase in cell to cell spread. Infect. Immun ., 1992, 60 , 219-230.
[19] ROUQUETTE C., TASCON R., PELLEGRINI E., BOLLA J.M., RIPIO M.T., VAZQUEZ-BOLAND J.A., BERCHE P. — Identification of a ClpC ATPase required for stress tolerance and in vivo survival of Listeria monocytogenes.
Mol. Microbiol. , 1996, 21 , 977-988.
[20] ROUQUETTE C., DE CHASTELLIER C., NAIR S., BERCHE P. — The ClpC ATPase of Listeria monocytogenes is a general stress protein required for virulence and promoting early bacterial escape from the phagosome of macrophages. Mol. Microbiol. , 1998, 27 , 1235-1245.
[21] NAIR S., DERRÉ I., MSADEK T., GAILLOT O., BERCHE P. — CtsR is a repressor of the class III heat shock genes in the human pathogen Listeria monocytogenes. Mol. Microbiol., 2000 , 34 , 800-811.
[22] GAILLOT O., PELLEGRINI E., BREGENHOLT S., NAIR S., BERCHE P. — The serine-protease ClpP is essential for the intracellular parasitism of the pathogen Listeria monocytogenes . Mol. Microbiol., 2000, 35 , 1286-1294.
[23] NAIR S., MILOHANIC E., BERCHE P. —The ClpC ATPase is required for cell adhesion and invasion of Listeria monocytogenes . Infect. Immun., 2000, 68 , 7061-7068 [24] TILNEY L.G., CONNELLY P.S., PORTNOY D.A. — Actin filament nucleation by the bacterial pathogen, Listeria monocytogenes. J. Cell Biol., 1990, 111 , 2979-2988.
[25] PRON B., BOUMAILA C., JAUBERT F., SARNACKI S., MONNET J.P., BERCHE P., GAILLARD J.L. — Comprehensive study of the intestinal stage of listeriosis in a rat ligated ileal loop system. Infect.
Immun., 1998, 66 , 747-755.
[26] PRON B., BOUMAILA C., JAUBERT F., BERCHE P., MILON G., GAILLARD J.L. — Dendritic cells are early cellular targets of Listeria monocytogenes after intestinal delivery and are involved in bacterial spread in the host, Cell. Microbiol. , 2001, 3 , 1-11 [27] JOIN-LAMBERT O., EZINE S., LE MONNIER A., JAUBERT F., OKABE M., BERCHE P., KAYAL S. — Listeria monocytogenes-infected bone-marrow myeloid cells promote bacterial invasion of the central nervous system. Cell. Microbiol., 2005 , 7 , 167-180.
[28] GAILLARD J.L., JAUBERT F., BERCHE P. — The inlAB locus mediates the entry of Listeria monocytogenes into hepatocytes in vivo . J. Exp. Med. , 1996, 183 , 359-369.
[29] BERCHE P.— Bacteremia is required for invasion of the murine central nervous system by Listeria monocytogenes. Microb. Pathog. , 1995, 18 , 323-336.
[30] DREVETS D.A. — Dissemination of Listeria monocytogenes by infected phagocytes.
Infect.
Immmun ., 1999, 67 , 3512-3517.
[31] DREVETS D.A., DILLON M.J., SCHAWANG J.S., VAN ROOIJEN N., EHRCHEN J., SUN DERKOTTER C., LEENEN P.J. — The Ly-6C (high) monocyte subpopulation transports Listeria monocytogenes into the brain during systemic infection of mice . J. Immunol., 2004 , 172, 4418-4424.
[32] KAYAL S., LILIENBAUM A., POYART C., MEMET S., ISRAEL A., BERCHE P. — Listeriolysin O-dependent activation of endothelial cells during infection with Listeria monocytogenes :
upregulation of adhesion molecules, chemokines, and transcription factor NF-kB. Mol. Microbiol. , 1999, 31 , 709-722.
[33] KAYAL S., LILIENBAUM A., LOIN-LAMBERT O., LI X., ISRAEL A., BERCHE P. — Listeriolysin O secreted by Listeria monocytogenes induces NFkB activation by activating the I-kB kinase complex, Mol. Microbiol. , 2002, 44 , 1407-1419 [34] BERCHE P. — Infections materno-fétales à Listeria monocytogenes, in ‘‘
Bactéries, Champignons et parasites transmissibles de la mère à l’enfant ’’, F Denis, Ed. John Libbey, 2002, 91-106.
DISCUSSION
M. Léon LE MINOR
Quelle est l’incidence des Listeria dans les avortements spontanés ? Quels sont les marqueurs bactériens utilisables dans les enquêtes épidémiologiques ? Comment cette surveillance est-elle organisée en France ? Y a-t-il des aliments qui sont plus fréquemment en cause dans les infections à Listeria ? Y a-t-il une plus grande concentration de ces bactéries dans la croûte des fromages ?
On ne connaît pas avec certitude l’incidence de
L. monocytogenes dans les avortements spontanés. Cependant, il est très probable que, parmi les causes infectieuses possibles, cette bactérie représente une part non négligeable d’avortements spontanés. Les marqueurs bactériens actuellement utilisables dans les enquêtes épidémiologiques sont tout d’abord, une fois que l’espèce L. monocytogenes est clairement identifiée, la sérotypie et la lysotypie (sensibilité aux phages). Ces méthodes sont actuellement supplantées par l’analyse des empreintes génétiques (habituellement réalisés par électrophorèse en champs pulsés). Il faut savoir que seulement quelques pays en Europe (France, Belgique, Royaume-Uni…) possèdent des centres de référence qui collectent l’ensemble des souches. Les épidémies de listériose sont la plupart du temps dues à la diffusion large sur l’ensemble d’un pays d’un produit commercialisé contaminé. Les centres de référence qui
réalisent l’étude systématique des empreintes génétiques des souches isolées de patients peuvent ainsi identifier une épidémie en repérant des souches identiques dans des endroits parfois très éloignés. En effet, on sait que les souches sont naturellement très hétérogènes, et que la probabilité d’avoir des souches identiques par hasard reste très faible. Les pays qui ont des centres de référence déclarent plus d’épidémies que les autres, mais alertent plus rapidement les pouvoirs publiques et donc limitent fortement ces épidémies. En France, le centre de référence est localisé à l’Institut Pasteur de Paris. Les aliments les plus fréquemment en cause sont les fromages à pâte molle, les charcuteries (rillettes, langue de porc, pâté…), les biftecks hachés, certains poissons fumés (saumon…), certains végétaux (salades, soja…), pour ne citer que les principaux. Chez l’animal, il faut rappeler le rôle des ensilages massivement contaminés par la bactérie dans les zoonoses. La prévention de la listériose est difficile car L. monocytogenes est un germe saprophyte du sol et contamine donc en faibles quantités de nombreux aliments.
Nous sommes donc exposés en permanence à une faible contamination par cette bacté- rie. Malgré un contrôle très rigoureux des processus industriels de préparation des aliments, il peut arriver tout à fait rarement que certains aliments soient contaminés, notamment du fait de la capacité de la bactérie à se multiplier à 4° C. Enfin, la cuisson des aliments reste le meilleur moyen pour prévenir la listériose, la bactérie étant facilement détruite par la chaleur. Quant à la croûte du fromage, il semble qu’effectivement on puisse trouver une plus forte quantité de bactéries dans la croûte que dans la pâte dans certains fromages contaminés.
Mme Jeanne BRUGÈRE-PICOUX
Vous avez souligné l’importance de la bactériémie dans la pathogénie de la listériose. Or, en médecine vétérinaire, il avait été démontré une corrélation entre l’éruption des bourgeons dentaires chez le mouton et l’apparition des troubles nerveux ultérieurs (essentiellement une paralysie faciale). La voie nerveuse peut-elle être retenue dans ce cas ?
En pathologie humaine, il semble clair que l’infection est la conséquence de l’absorption d’aliments contaminés, induisant une bactériémie exposant le placenta et le système nerveux central. Les formes encéphalitiques pures entraînant des rhombencéphalites sans méningite chez certains animaux comme les ovins, ont fait évoquer une voie nerveuse rétrograde, où les bactéries gagneraient le tronc cérébral notamment par les filets du nerf olfactif. Ceci est possible mais très peu de travaux expérimentaux ont pu corroborer cette hypothèse. Il faudrait des études anatomopathologiques fines, visualisant les bactéries dans les filets nerveux des nerfs crâniens pour affirmer cette voie. Une des difficultés est qu’il est difficile d’éliminer une éventuelle bactériémie d’origine alimentaire chez les animaux vivants dans des conditions précaires de stabulation et éventuellement infectés par de multiples voies.
M. Jean-Paul GIROUD
Quels sont les antibiotiques actuels les plus actifs dans les atteintes méningées vis-à-vis de Listeria monocytogenes ?
La bactérie reste sensible à la plupart des antibiotiques actifs sur les bacilles à Gram positif. Les antibiotiques les plus actifs sont la pénicilline G, l’ampicilline (et amoxicilline), le triméthoprime-sulfaméthoxazole, les aminosides tels que la gentamicine, la
rifampicine, les tétracyclines et même le chloramphénicol. Il existe quelques souches résistantes aux tétracyclines et exceptionnellement des souches exprimant plusieurs résistances. Un point important à considérer dans la stratégie thérapeutique : L. monocytogenes est une bactérie intracellulaire nécessitant une bonne diffusion tissulaire des antibiotiques. Paradoxalement, l’ampicilline pénètre bien dans le cytoplasme des cellules où les bactéries se multiplient, et peu dans les phagosomes. C’est l’antibiotique de choix pour le traitement, habituellement utilisé en association à la gentamicine. L’alternative est l’association du triméthoprime-sulfaméthoxazole à la gentamicine.
M. Roland ROSSET
Des problèmes de nombre ! Comment expliquer un nombre si faible de listeriose en France (225) alors que nous consommons tous des Listeria tous les jours ? Dose infectante : dans vos expériences, vous nous montrez la nécessité d’injecter à l’animal des 109 de germes. Quid pour l’homme ? Quelle est la dose minimum nécessaire ? Pouvez-vous préciser l’importance du système immunitaire dans la résistance ou la susceptibilité du consommateur ?
L. monocytogenes est une bactérie du sol qui nous contamine accidentellement. Elle n’a pas été « éduquée » au cours de l’Évolution à traverser le tube digestif, mais elle a la capacité de se multiplier dans les cellules du fait de facteurs de virulence sélectionnées pour survivre aux prédateurs naturels de cette bactérie dans le sol (amibes…). La bactérie n’a donc que peu de facteurs lui permettant de franchir le tube digestif, à la différence de bactéries hautement adaptées aux animaux et à l’homme depuis le néolithique, telles que les salmonelles, qui ont parfois une dose infectante souvent très faible de l’ordre de 103 bactéries. Les salmonelloses sont beaucoup plus fréquentes (et heureusement beaucoup moins graves) que la listériose, pouvant atteindre dans notre pays plus de 20 000 cas déclarés par an comparativement aux 200 cas environ de listériose déclarés chaque année.
Malgré une exposition alimentaire à L. monocytogenes très fréquente, itérative et à faible dose, le nombre de cas reste très faible car la bactérie franchit difficilement la barrière digestive et les rares bactéries qui passent sont contrôlées par le système immunitaire.
Après absorption d’une forte dose de bactéries, rares vont être ceux qui vont développer une listériose. Seuls vont payer tribut à la maladie les sujets fragiles, femmes enceintes, patients immunodéprimés, et peut-être patients présentant une sensibilité génétique particulière à la bactérie. La dose infectante n’est pas connue avec certitude chez l’homme mais lors des épidémies documentées, les patients auraient absorbé des doses oscillant de 105 à 109 bactéries.
M. Henri LECLERC
Pourquoi la listériolysine qui détruit la membrane du phagocyte ne détruit-elle pas la membrane cytoplasmique ? La listériolysine est-elle une toxine ou une structure apparentée au système de secrétion des bacilles à gram négatif, permettant à la protéine d’agir ? Les listeria sont des bactéries de l’environnement. Comment ont-elles fait pour surmonter à la fois le déficit nutritionnel et les différents stress rencontrés dans l’environnement (choc oxydatif, thermique, osmotique, etc.) ? Est-ce le fait de la réponse stringente avec le psppGpp et de la réponse générale aux stress contrôlée par le régulateur RpO5 (facteur sigma S) ?
Il est démontré que, lors de l’accès au cytoplasme, la bactérie cesse de produire la listériolysine O qui devient indétectable. Tout se passe comme si le changement
d’environnement (acide, carencé et hostile dans le phagosome, neutre et favorable à la croissance dans le cytoplasme) induisait un arrêt de la transcription du gène codant cette toxine, transcription dépendant de l’activateur transcriptionnel PrfA. La listériolysine O est une toxine qui, par polymérisation, forme des pores dans la membrane des phagosomes, entraînant la lyse cellulaire. La vie dans l’environnement d’une bactérie saprophyte n’est pas de tout repos. En effet, il faut garder à l’esprit que les bactéries sont presque continuellement soumises à des variations de température, au rayonnement ultraviolet du soleil, aux prédateurs et compétiteurs naturels qui les entourent, aux carences nutritionnelles chroniques, pour n’en citer que quelques-uns. La survie dans les conditions hostiles représente en fait l’essentiel du temps d’une bactérie saprophyte. Au cours de cette dure existence, les bons moments, ceux de la croissance exponentielle liée à des conditions favorables avec des nutriments en abondance, sont tout à fait rares. Pour toutes ces raisons, Listeria a, comme d’autres bactéries saprophytes, développé des systèmes sophistiqués de réponse aux stress thermiques, oxydatifs, nutritionnels, chimiques, osmotiques… Ces systèmes lui sont d’ailleurs utiles pour survivre dans les cellules comme nous avons pu le démontrer récemment.
M. Jean-François BACH
Parmi les états d’immunodépression prédisposant à la survenue d’une listériose chronique, existe-t-il des éléments communs qui pourraient orienter vers un mécanisme immunitaire clef dans la défense immunitaire contre la bactérie ?
On sait qu’il existe une immunité innée contre
L. monocytogenes associée à une réponse inflammatoire rapide mobilisant des polynucléaires neutrophiles pour lyser les cellules infectées, exposant ainsi les bactéries à ces polynucléaires hautement bactéricides.
Conjointement, il existe une immunité adaptative liée à la production de lymphocytes CD8 cytotoxiques, qui vont agir dans le même sens. Près des deux tiers des patients présentant une listériose ont des facteurs de risque : femmes enceintes au 3ème trimestre de la grossesse, patients sous chimiothérapie pour cancer ou pour greffe, sidéens, patients avec hémochromatose, diabétiques… Les deux facteurs essentiels de prédisposition apparaissent donc être la neutropénie et un affaiblissement de l’immunité cellulaire. Quant aux malades apparemment sains qui développent une listériose, il est possible qu’ils présentent une prédisposition génétique les rendant plus sensibles à cette bactérie, qu’il s’agisse de l’altération de l’immunité innée ou acquise contre les antigènes bactériens.
M. Jacques-Louis BINET
Existe-t-il une traduction clinique de la listériose, monocytose ou neutropénie ? Quelle est la sensibilité de la listeria à la température ? Peut-elle se développer à + 4° ?
La bactérie a été découverte en 1926 au cours d’une épidémie dans un élevage de lapins qui présentaient une forte monocytose. C’est de là que vient le nom d’espèce monocytogenes. Cependant, la monocytose peut varier selon l’espèce animale. Chez l’homme, elle est inconstante et, si le patient n’est pas neutropénique avant infection, on peut même observer une hyperleucocytose avec de nombreux polynucléaires. La cuisson des aliments est la base de la prévention de la listériose. Très bien adapté à l’environnement, la bactérie est capable de croître à basse température, en particulier à 4° C, ce qui est fondamental pour l’épidémiologie de la maladie. En effet, les aliments soumis à un processus indus-
triel, sont souvent stockés pendant un temps relativement long dans des réfrigérateurs et les bactéries peuvent se multiplier lentement à basse température, atteignant sans altérer le goût des aliments des taux parfois très élevés, de l’ordre de 100.000 à 10 ou 100 millions de bactéries par gramme d’aliments.
M. Pierre GODEAU
Où se situe le point d’impact de l’immunodépression permettant le développement et la diffusion de l’infection à L. monocytogenes ?
La stratégie des bactéries est la quête d’un sanctuaire cellulaire, qui puisse les protèger des polynucléaires qui les détruisent instantanément. Les bactéries peuvent survivre et se multiplier dans la plupart des cellules de l’organisme, y compris les macrophages, sans forcément les lyser rapidement. Les monocytes et les cellules dendritiques peuvent même véhiculer les bactéries dans la circulation sanguine. La réponse de l’immunité innée et adaptative vise à détruire les cellules infectées exprimant des antigènes bactériens à leur surface, pour exposer les bactéries aux polynucléaires. Le point d’impact de l’immunodépression est donc d’affaiblir la capacité cytolytique des polynucléaires neutrophiles et des lymphocytes, rendant l’hôte infecté incapable de contrôler la croissance intracellulaire de Listeria .
Francois DENIS
Que sait-on de l’immunité muqueuse antilistérienne au niveau intestinal ?
Peu de choses. Il existe une sécrétion locale d’IgA contre les bactéries et leurs toxines, dont le rôle pourrait être d’opsoniser les bactéries, et donc de protéger un peu plus la muqueuse intestinale que Listeria a déjà du mal à franchir. C’est un mécanisme supplé- mentaire pouvant expliquer la rareté de la maladie par rapport à la fréquence de l’exposition.
M. Gilles CRÉPIN
La listeria est particulièrement redoutée pour les femmes enceintes au 3è trimestre où le placenta est altéré et beaucoup plus perméable aux bactéries. L’ampicilline à fortes doses (minimum 3g/24h) est utilisée. Les données bactériologiques actuelles confirment-elles l’absence de résistance à l’ampicilline ? Par ailleurs, existe-t-il une différence de sensibilité en fonction de la souche de Listeria monocytogenes ?
Aucune souche résistante à l’ampicilline n’a jusqu’ici été décrite. Cet antibiotique reste le traitement de choix de la listériose. Toutes les souches semblent avoir le même niveau de sensibilité à l’ampicilline. Les différences d’efficacité du traitement en fonction des patients sont en fait surtout le reflet de l’ampleur des dégâts neurologiques ou placentaires au moment du début du traitement. La gravité de la maladie est liée au fait que lorsque le diagnostic est posé, par exemple quand le patient développe une rhombencéphalite, les lésions sont déjà très importantes.
M. Marc GIRARD
Vous avez mentionné, à côté de la forme classique de la listériose (infection fébrile à complications méningo-encéphalitiques et avortements), l’existence de toxi-infections alimentaires avec diarrhée et fièvre. Que sait-on sur cette nouvelle forme de la maladie ?
Est-elle la simple conséquence de contaminations massives ou est-elle liée à l’émergence d’un variant bactérien particulier ? Signe-t-elle par exemple un tropisme accru de la bactérie pour l’entérocyte ? La listériolysine O est-elle la seule toxine impliquée dans ces infections ?
La plupart du temps, la listériose est une maladie fébrile qui n’entraîne pas de diarrhée.
De plus, le temps d’incubation peut être de plusieurs jours à plusieurs semaines. Cependant, on a récemment décrit plusieurs épidémies de listériose, notamment en Italie et aux États-Unis, chez des sujets sains présentant une toxi-infection alimentaire fébrile, avec diarrhée aiguë. Ce que nous montrent ces épidémies, c’est que l’absorption de doses très fortes de bactéries peut entraîner en quelques heures une gastro-entérite fébrile. La souche n’avait rien de particulier et ne semble pas un variant nouveau de la bactérie. C’est plutôt le mode de contamination qui est à incriminer. De plus, ces épidémies mettent en exergue la méconnaissance possible de cette cause de toxi-infection alimentaire dans le passé.
* Faculté de médecine René Descartes Paris 5, 15 rue de l’École de Médecine, 75015 Paris. Tirés-à-part : Professeur Patrick BERCHE, même adresse. Article reçu le 15 novembre 2004, accepté le 24 janvier 2005.
Bull. Acad. Natle Méd., 2005, 189, no 3, 507-521, séance du 15 mars 2005