
Résumé
Pasteur a mis la vaccination sur un pied empirique et expérimental pendant les années 1880, et le développement des vaccins a procédé lentement jusqu’à la deuxième guerre mondiale. Puis les vaccins vivants contre maladies bactériennes et virales ont été atténués par le passage chez les animaux. Les microbes ainsi tués ont été inactivés sans pourtant détruire leur pouvoir immunogène. De plus, la connaissance des toxines bactériennes et polyosides capsulaires a permis le développement de nouveaux vaccins pour plusieurs maladies épidé- miques. Au début du troisième siècle de la vaccination, les méthodes classiques obtiennent toujours des nouveaux vaccins, mais la biologie moléculaire et la génie génétique ont commencé à influer sur leur développement. De plus, pour la première fois l’immunologie fondamentale apporte une contribution au domaine de la vaccinologie. Donc, les tendances actuelles sont les suivantes : le réassortiment des génomes segmentés, les souches atténuées recombinées avec les gènes des pathogènes, les vecteurs portant des gènes étrangers, les particules réplication-défectives, les plasmides d’ADN, et la vaccinologie inverse, entre autres. Aussi, les nouvelles méthodes de livraison hors de l’injection seront employées et des nouveaux adjuvants rajoutés aux vaccins pour stimuler les réponses immunes spécifiques. L’avenir de la vaccination est donc prometteur.
Summary
Pasteur put vaccination on an empiric and experimental basis during the 1880s, and vaccine development proceeded slowly until the second World War. During this period live vaccines against bacterial and viral diseases were developed by attenuation through passage in animals and killed microbes were inactivated without destroying their immunogenicity. Moreover, knowledge of bacterial toxins and polysaccharides permitted the development of new vaccines for several epidemic diseases. At the beginning of the third century of vaccination, classical methods are still providing new vaccines, but molecular biology and genetic engineering have begun to influence vaccine development. In addition, for the first time basic immunology is contributing to the domain of vaccinology. Thus, the current trends in vaccine development are as follows : reassortment of segmented genomes, attenuated strains recombined with genes from pathogens, vectors carrying foreign genes, replication-defective particles, DNA plasmids, and reverse vaccinology, among others. Also, new methods of vaccine delivery besides injection will be used and new adjuvants will be added to vaccines in order to stimulate specific responses. The future of vaccination is promising.
Comme nous le savons tous, la vaccination, à distinguer de la variolisation, a vu le jour grâce aux observations de Jenner sur la protection conférée par la vaccine. Ce fut le premier des vaccins atténués. Mais Pasteur fut le premier à placer la vaccination sur le plan expérimental en développant des méthodes pour inactiver les virus et les bactéries. La dichotomie entre les vaccins atténués et inactivés reste utile aujourd’hui, même si, comme nous le verrons, les nouvelles stratégies vaccinales rendent la distinction moins claire.
Le tableau 1 cite certaines de ces nouvelles stratégies. Dans le temps qui m’est imparti, je n’aurai le temps d’en aborder que six.
TABLEAU 1. — Stratégies les plus récentes pour le développement de vaccins Réassortiment de segments de génomes de RNA Agents réplicants recombinés avec des gènes de pathogènes.
Vecteurs Particules Réplication — défectives Plasmides ADN Vaccinologie inverse Protéines recombinantes génétiquement inactivées Protéines de fusion Administration de gènes par des bactéries invasives Combinaison vaccinales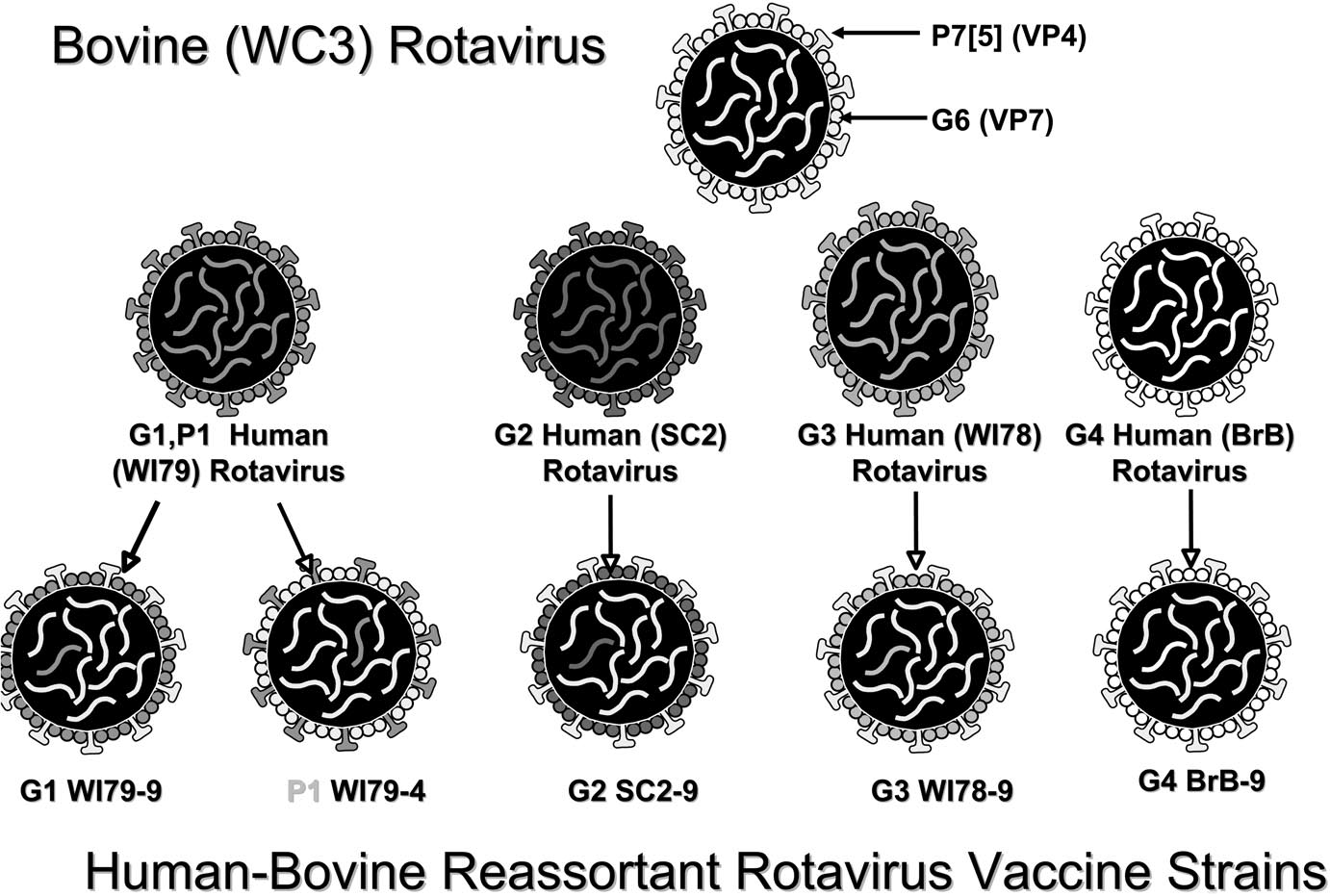
FIG. 1 Le réassortiment est un procédé qui peut être utilisé avec les virus à ARN qui ont des génomes segmentés. Dans mon ancien laboratoire à Philadelphie, le réassortiment a été utilisé pour créer le virus pentavalent contre le rotavirus qui est aujourd’hui autorisé [1]. Les rotavirus ont onze segments d’ARN double brin, chacun codant pour des protéines virales. Parmi ces dernières, on trouve les protéines G et les protéines P qui portent des épitopes neutralisants et qui sont situées à la surface du virus (figure 1). Nous avons isolé un rotavirus bovin, appelé WC-3, qui est naturellement atténué pour l’être humain. Bien que le virus bovin ait conféré une certaine protection contre les infections à rotavirus humain, il a été sensiblement amélioré par le placement de protéines G et P issues de virus humains à sa surface. Cela a été réalisé par co-infection du virus bovin et de virus humains en culture cellulaire avec sélection des réassortants portant dix segments d’ARN du virus bovin et un segment d’ARN d’un virus humain. Cinq de ces réassortants, chacun portant une protéine de rotavirus humain différente à sa surface, ont été combinés pour fabriquer le vaccin, qui est très efficace. Le vaccin est administré par voie orale en trois doses et il est efficace à plus de 95 % contre les infections graves à rotavirus.
Un autre vaccin a été développé par réassortiment, il s’agit du nouveau vaccin vivant atténué contre la grippe. Ce vaccin est administré par insufflation intranasale plutôt que par injection. Mais le principal intérêt du vaccin vivant n’est pas la commodité d’éviter les injections, mais la preuve que l’immunité conférée est plus large que celle obtenue après l’administration du vaccin inactivé. Le tableau 2 et
[2-4] montrent que même lorsque la souche de virus sauvage circulante a changé par rapport à la souche contenue dans le vaccin, une efficacité élevée a été maintenue chez l’enfant. En outre, lorsqu’un grand nombre d’enfants d’une communauté ont été vaccinés, les visites médicales ont diminué d’environ 60-70 % aussi bien parmi les enfants que parmi leurs contacts adultes, ce qui témoigne d’une immunité de groupe.
TABLEAU 2. — Protection hétérovariante par le vaccin grippal intranasal chez l’enfant [2, 3, 4] Saison Souche vaccinale Souche circulante Efficacité % (I.C.) 96-97 A/Wuhan (H3N2) A/Wuhan 96 (90-99) 96-97 A/Wuhan (H3N2) A/Sydney 86 (77-93) 00-01 A/Sydney (H3N2) A/Panama 89 (67-99) 00-01 A/Beijing (H1N1) A/New Caled.
84 (-11-96)*
00-01 B/Beijing B/Sichuan 66 (9-87)*
Le virus respiratoire syncytial est le virus respiratoire le plus important chez le nourrisson, mais les efforts pour développer un vaccin ont été lents. Un laboratoire des National Institutes of Health des Etats-Unis a atténué un virus parainfluenza de type 3 (responsable d’infections respiratoires chez le nourrisson), puis a inséré un gène du VRS codant pour la protéine de fusion (F). Ce virus recombinant a ensuite été utilisé pour vacciner des singes, qui ont ensuite été exposés au VRS sauvage.
Comme le montre le tableau 3 et [5] alors que les singes témoins ont excrété beaucoup de virus après l’exposition, les singes vaccinés ont été complètement protégés. Ainsi, la recombinaison de deux virus est une stratégie intéressante.
TABLEAU 3. — Les singes vaccinés avec du PIV3/VRS bovin/ humain étaient efficacement protégés d’un challenge avec du VRS.[5] Moyenne des titres maximaux (log10 UFP/ml fi SE Virus vaccinant Postchallenge NP BAL <1.2 fi 0.4 <1.2 fi 0.3 b/h PIV3/VRS F2 Placebo 4.3 fi 0.3 5.7 fi 0.3 Une stratégie largement utilisée pour les vaccins expérimentaux recourt à des vecteurs, qui sont définis comme des véhicules non pathogènes dans lesquels des gènes issus de pathogènes ont été insérés, exprimant les protéines correspondantes.
Les poxvirus, les adénovirus et le Bacille Calmette-Guérin sont des exemples de vecteurs. Un autre exemple est la souche de fièvre jaune atténuée 17D, qui peut être utilisée comme vecteur pour d’autres flavivirus tels que le virus de la dengue ou le virus du Nil occidental. Pour construire un vaccin contre la dengue, les gènes de l’enveloppe du virus de la fièvre jaune ont été excisés et les gènes correspondants de chacun des quatre sérotypes du virus de la dengue ont été insérés, donnant un vaccin quadrivalent. Ce vaccin expérimental contre la dengue est immunogène et il est en cours d’essais de phase III pour déterminer son efficacité. Un autre exemple de l’utilisation de vecteurs provient de nos travaux pour tenter de développer un vaccin contre le cytomégalovirus humain. La principale cible de l’immunité cellulaire contre le CMV est l’antigène de la matrice pp65. Le gène codant pour pp65 a été inséré dans un virus canarypox, qui est inoffensif pour l’être humain. L’injection de deux doses du canarypox-pp65 a induit de fortes réponses des cellules T cytotoxiques, comme le montre la figure 2 et [6].
utilisée comme vecteur pour d’autres flavivirus tels que le virus de la dengue ou le virus du Nil occidental. Pour construire un vaccin contre la dengue, les gènes de l’enveloppe du virus de la fièvre jaune ont été excisés et les gènes correspondants de chacun des quatre sérotypes du virus de la dengue ont été insérés, donnant un vaccin quadrivalent. Ce vaccin expérimental contre la dengue est immunogène et il est en cours d’essais de phase III pour déterminer son efficacité. Un autre exemple de l’utilisation de vecteurs provient de nos travaux pour tenter de développer un vaccin contre le cytomégalovirus humain. La principale cible de l’immunité cellulaire contre le CMV est l’antigène de la matrice pp65. Le gène codant pour pp65 a été inséré dans un virus canarypox, qui est inoffensif pour l’être humain. L’injection de deux doses du canarypox-pp65 a induit de fortes réponses des cellules T cytotoxiques, comme le montre la figure 2 et [6].
FIG. 2. — Réponse CMV CTL au vecteur Canarypox-pp65[6 Une quatrième stratégie nouvelle est celle des particules pseudo-virales. Essentiellement, cette stratégie découle de la capacité de certaines protéines virales structurelles à s’auto-assembler si elles sont produites par des lignées cellulaires dans lesquelles les gènes codant pour les protéines ont été insérés. L’exemple le plus connu est celui du nouveau vaccin contre les papillomavirus humains responsables de cancers du col utérin. Il est fabriqué à partir de la production d’une protéine appelée L-1 par des cellules ovariennes de hamster chinois ou baculovirus. Les particules ainsi produites ressemblent à des virus HPV infectieux, mais elles sont dépourvues d’ADN et sont incapables de se répliquer. En revanche, elles sont très immunogènes et se sont révélées efficaces vis-à-vis des transformations cancéreuses des cellules de col de l’utérus dues au HPV, ainsi que des verrues génitales. Des particules pseudovirales ont été développées pour de nombreux autres agents, notamment le virus de la grippe, le VIH et le virus Ebola, responsable d’une fièvre hémorragique. Les études chez l’animal montrent une efficacité élevée de ces vaccins.
Les découvertes scientifiques sont souvent faites par un heureux hasard et tel fut le cas de la découverte que les plasmides d’ADN contenant des gènes codant pour des antigènes pouvaient exprimer les protéines correspondantes après injection et induire des réponses immunitaires. Cette découverte fortuite a été largement appliquée aux vaccins expérimentaux. Même si, au début, on a trouvé que de grandes quantités d’ADN devaient être injectées pour réussir à immuniser, on a ensuite trouvé (tableau 4) que les plasmides d’ADN sont d’excellentes amorces pour le système immunitaire et que, dans le cas des animaux nouveau-nés, l’amorçage pouvait surmonter l’inhibition de la réponse immunitaire par les anticorps maternels. Par ailleurs, le système immunitaire néonatal a été orienté vers une réponse Th1 plutôt que la réponse Th2 habituelle. En outre, les plasmides d’ADN ont induit une forte mémoire immunologique.
TABLEAU 4. — Vaccination par les plasmides de ADN.[6] ➣ Induit un priming du système immunitaire ➣ Induit un priming de l’immunité néonatale pour pallier la suppression des anticorps maternels.
➣ Oriente le système immunitaire néonatal vers une différentiation TH1 ➣ Induit une mémoire immunologique forte TABLEAU 5. — Vaccinologie inverse [7] 1. Séquençage génomique complet du pathogène 2. Insertion des gènes dans. E. coli 3. Expression des protéines par
E. coli.
4. Immunisation des souris avec les protéines 5. Immunisation des souris avec les protéines exprimées.
6. Tester les serums pour l’activité bactéricide 7. Déterminer la conservation des antigènes parmi les souches de pathogène Le problème de la nécessité de quantités importantes de plasmides d’ADN semble avoir été récemment résolu par l’utilisation d’adjuvants ou par électroporation (application d’un faible courant électrique au site d’injection). Actuellement, les plasmides d’ADN codant pour des antigènes jouent un rôle majeur dans les tentatives pour développer de nouveaux vaccins sous forme de primo-vaccinations suivies de rappels par des protéines ou des vecteurs portant les mêmes gènes.
La dernière stratégie discutée est celle de la vaccinologie inverse (tableau 5) et[7]. Ce concept signifie essentiellement que l’on commence par le génome d’une bactérie pathogène et, une fois sa séquence connue, on déduit quels ‘‘ cadres de lecture ouverts ’’ codent pour les protéines qui sont situées à la surface d’un organisme ou
qui sont sécrétées par l’organisme. Elles seront probablement utiles comme immunogènes. Les cadres de lecture ouverts (gènes) sont chacun insérés dans E. coli pour la production des protéines et les protéines sont ensuite utilisées pour immuniser des souris. Le sérum des souris est examiné à la recherche d’une activité bactéricide. Si un sérum est bactéricide vis-à-vis d’un vaste éventail de souches bactériennes, il devient candidat à la poursuite du développement. Cette technique a notamment été appliquée aux méningocoques du groupe B, avec des résultats prometteurs.
Le développement de nombreux vaccins nouveaux, s’ils sont tous administrés par injection, pose un problème d’acceptabilité et aussi d’interférence éventuelle entre antigènes. Un développement important dans l’application des vaccins est donc l’invention d’une technologie pour administrer ceux-ci par voie transcutanée.
L’administration transcutanée repose sur la nécessite de rendre l’épiderme plus poreux à l’absorption et sur la présence de nombreuses cellules de Langerhans dans le derme, qui capturent les antigènes et les transportent dans les ganglions lymphatiques adjacents. L’absorption des antigènes d’un patch appliqué sur la peau peut être augmentée par l’adjonction d’adjuvants au patch ou par l’application de faibles courants électriques au site d’application du patch.
Une méthode encore plus simple consiste à utiliser des microaiguilles de 150 microns de long pour transpercer l’épiderme mais de ne pas entrer dans le tissu sous-cutané. Des microaiguilles sont aujourd’hui au point et seront certainement autorisées. Des études conduites par Sanofi Pasteur ont montré que l’injection du vaccin antigrippal à l’aide d’une microaiguille produit des réponses d’anticorps supérieures à celles obtenues après l’injection intramusculaire.
CONCLUSION
Il existe un éventail de nouvelles stratégies pour développer des vaccins sur la base des découvertes de la biologie moléculaire. De nombreuses cibles demeurent, comme le VIH, la tuberculose et le paludisme, mais nous pouvons compter sur un âge d’or de la vaccination à condition que la société ait la sagesse de les utiliser car mieux vaut prévenir que guérir [8].
REMERCIEMENTS
Je voudrais remercier M. Keith Veitch de son aide indispensable pour le texte français.
BIBLIOGRAPHIE [1] CLARK H.F., OFFIT P.A., PLOTKIN S.A., HEATON P.M. — The new pentavalent rotavirus vaccine composed of bovine (strain WC3)-human rotavirus reassortants. Ped. Infect. Dis. J., 2006, 25 , 577-583.
[2] BELSHE R., LEE M.S., WALKER R.E. ET al. — Safety, immunogenicity and efficacy of intranasal, live attenuated influenza vaccine.
Expert Rev. Vaccines, 2004, 3 , 643-54.
[3] LEE M.S., MAHMOOD K., ADHIKARY L., AUGUST M.J., CORDOVA J., CHO I., KEMBLE G., REISINGER K., WALKER R.E., MENDLEMAN P.M. — Pediatr. Infect. Dis. J., 2004, 23 , 852-6.
[4] HALLORAN M.E., LONGINI IM J.R., GAGLANI M.J., PIEDRA P.A., CHU H., HERSCHLER G.B., GLEZEN W.P. — Estimating efficacy of trivalent, cold-adapted, influenza virus vaccine (CAIV-T) against influenza A (H1N1) and B using surveillance cultures. Am. J. Epidemiol ., 2003, 158 , 312-5.
[5] TANG R.S., MACPHAIL M., SCHICKLI J.H., KAUR J., ROBINSON C.L., LAWLOR H.A., GUZZETTA J.M., SPAETE R.R., HALLER A.A. — Parainfluenza virus type 3 expressing the native or soluble fusion (F) Protein of Respiratory Syncytial Virus (RSV) confers protection from RSV infection in African green monkeys. J. Virol . , 2004, 78, 20 , 11198-207.
[6] BERENCSI K., GYULAI Z., GONCZOL E., PINCUS S., COX W.I., MICHELSON S., KARI L., MERIC C., CADOZ M., ZAHRADNIK J., STARR S., STANLEY PLOTKIN S.A. — A Canarypox Vector-expressing cytomegalovirus phosphoprotein 65 (pp65) Induces Long-lasting Cytotoxic T Cell Responses in HCMV Seronegative Volunteers. J. Infect. Dis ., 2001, 183 , 1171-9.
[7] CAPECCHI B., SERRUTO D., ADU-BOBIE J., RAPPUOLI R., PIZZA M. — The genome revolution in vaccine research. Curr. Issues Mol. Biol., 2 004, 6 , 17-27.
[8] PLOTKIN S.A. — Vaccines : past, present, and future.
Nature Medicine, 2005, 11 , S5-S11.
DISCUSSION
M. Christian NEZELOF
Avec mes félicitations pour la clarté de votre exposé. Je voudrais avoir des commentaires supplémentaires sur l’utilisation des cellules de Langherans cutanés dont la répartition sur les téguments est inégale… Quels sont les adjuvants qui permettent l’étanchéité des kératinocytes ?
On utilise en général deux approches : soit rajouter au patch qui contient l’antigène un adjuvant comme la toxine labile de E. coli ou congénères que sont rendues moins toxiques par mutation, soit d’appliquer un petit courant électrique près du site d’injection ou de l’application du patch.
M. Jean-Jacques HAUW
Pouvez-vous nous indiquer, parmi les nouvelles stratégies que vous avez énumérées, celles qui sont les moins coûteuses et les moyens de réduire les prix de distribution ? Certaines d’entre elles sont-elles plus rapides à mettre en œuvre pour répondre à l’émergence d’une maladie infectieuse nouvelle ?
Vous touchez à un point d’importance capitale pour la dissémination de la vaccination partout dans le monde, et c’est ici le problème du coût des vaccins. Chaque nouveau vaccin coûte environs cinq cent millions de dollars à une société pharmaceutique, et les nouvelles stratégies seront toujours chères, parce que les frais importants sont ceux de contrôle de qualité. Les grandes sociétés pharmaceutiques du monde occidental espèrent
récupérer ces frais sur les bénéfices des pays riches, qui permettent un moindre prix dans les pays pauvres. Néanmoins, on ne peut pas vendre un produit à un prix inférieur aux coûts de la production. Donc, à mon avis, il y a deux solutions : le mécénat, comme le fait la Fondation Gates et la production par des sociétés localisées dans les pays hors de l’Europe ou des Etats-Unis, pays comme le Brésil, la Chine, l’Inde et autres.
M. Charles LAVERDANT
Peut-on expliquer pourquoi les vaccins plasmidiques induisent une mémoire immunologique aussi forte, voire définitive ?
La raison n’est pas entièrement connue, mais probablement il s’agit de la persistance des plasmides dans les cellules présentant des antigènes au niveau de la moelle, qui produisent la protéine immunisantes pour plus longtemps.
M. Roger NORDMANN
Pourriez-vous nous préciser quelles sont, à l’heure actuelle, les modalités de vaccination contre la tuberculose aux États-Unis ? Nous disposions, en France, du BCG administré à l’aide d’une bague. Beaucoup de pédiatres regrettent qu’il ne soit plus disponible sous cette forme. Quelle autre modalité préconisez-vous ? Y a-t-il de nouveaux vaccins en cours d’élaboration aux États-Unis ?
Le BCG n’est pas utilisé aux Etats-Unis pour la vaccination des nouveau-nés parce que l’incidence à cet âge est très basse. Bien que le BCG soit efficace pour la protection des enfants contre la tuberculose disséminée, il est peu efficace contre la tuberculose chez les adultes. Il y a maintenant plusieurs nouveaux vaccins en essai clinique. Pour la plupart, il y a deux approches suivies : soit insérer dans le génome du BCG des gènes de M.
tuberculosis qui codent pour des protéines soi-disant protectrices, soit d’atténuer M.
tuberculosis en enlevant les gènes de virulence.
M. Paul VERT
Les nouvelles stratégies de développement des vaccins permettraient-elles de contourner la contre-indication de vaccination des femmes enceintes par des virus vivants atténués ?
Oui, si nous pouvons exporter des Etats-Unis environ cinq cent mille avocats ! Plus sérieusement, le problème n’est pas qu’on ait une preuve que les vaccins sont dangereux pour la femme enceinte. Au contraire, plusieurs vaccins, comme ceux contre le tétanos et la grippe, sont indiqués chez la femme enceinte, mais il y a d’autres vaccins qui doivent être développés pour protéger le fœtus et le nouveau-né par les anticorps maternels. Ils sont bloqués pour des raisons médico-légales.
* Sanofi Pasteur, 4650 Wismer Road, Doylestown, PA 18902 USA Tirés à part : Professor Stanley A. PLOTKIN, même adresse
Bull. Acad. Natle Méd., 2008, 192, no 3, 511-519, séance du 4 mars 2008